圖說英國新冠變異株
凡人發了一篇很全麵的介紹[英國的新冠突變]的博文,給大家提供了交流互動的好機會。 言尤末盡,想到一圖至少頂10句 (A picture is worth a thousand words), 就來圖說一下這個之前稱B.1.1.7 現被命名為 'VUI-202012/01'的英國新冠變異株吧。
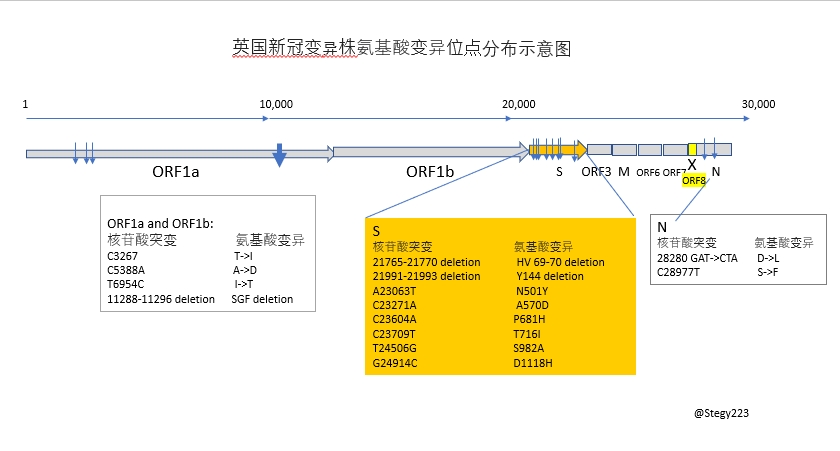
十七個氨基酸變異位點的分布:
決定每個氨基酸突變對蛋白質分子結構與功能的影響最主要有二點:
1) 突變發生在蛋白質分子三維結構的那個位點?
2) 什麽性質的突變?要談這點就得了解一下生物化學。20種氨基酸從每種的分子結構和化學反應性上講大體可分為5大組:
- 疏水性不與其它氨基酸分子作用的
- 親水性易與其它氨基酸分子形成氫鍵的
- 帶正電荷的,帶負電荷的
- 還有Pro 是唯一側鏈為環狀的。
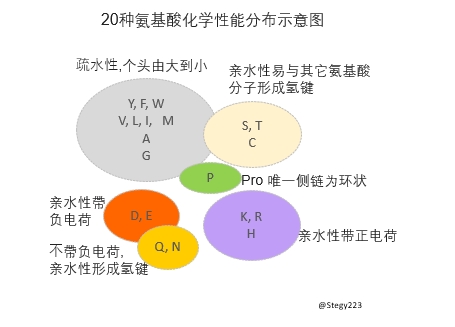
蛋白質分子比較能接受同類型的氨基酸變異(Conservative substitution) 。發生在蛋白質三維結構不同位點的不同類型的變異就有的可接受有的不太可接受。上述二點結合起來有些氨基酸突變對蛋白質分子的結構和功能可以沒什麽影響, 有些可以多多少少有一定程度影響,個別影響巨大。
眾知周知新冠刺突S蛋白是決定入侵細胞的關鍵蛋白,最終表現為傳染性的強弱。mRNA疫苗是基於對S蛋白與宿主細胞識別與結合的機理而量體裁衣設計的。所以目前關心的重點是S蛋白上的突變。就用個S蛋白三維結構示意圖來探討一下這一新冠變異株中S蛋白上的8個突變。
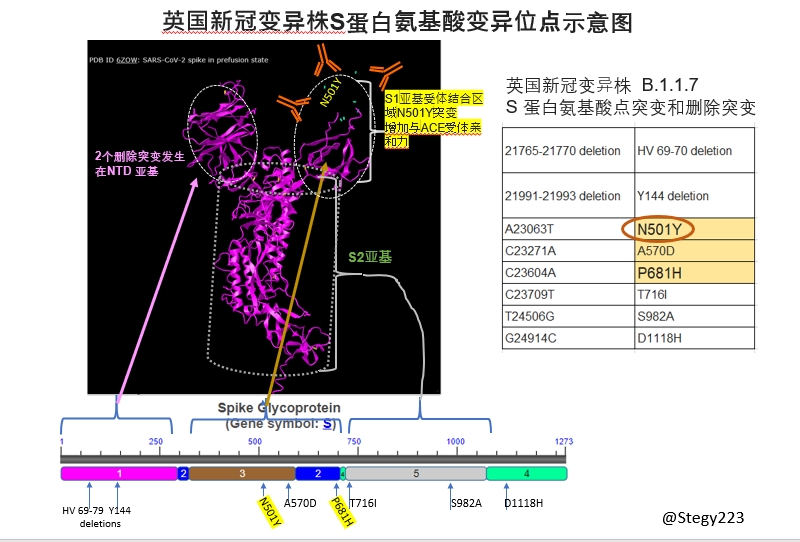
S蛋白在病毒膜外有三大塊。NTD亞基的功能不是太清楚,相比S1和S2亞基,進化上不是很保守,序列中突變多一些。S1亞基負責與宿主細胞膜上ACE2受體結合,S2亞基負責完成與細胞膜融合從而將病毒基因組RNA弄進宿主細胞。
2個刪除突變(HV69-70 deletion 和 Y144 deletion)發生在NTD亞基上,應該不會影響到爭對S1亞基產生的抗體。
S1亞基受體結合區域N501Y突變最受關注。之前在小白鼠動物模型實驗上已發現N501Y突變增強傳染性。N501是直接與ACE 受體結合的關鍵氨基酸之一。所以可以比較肯定地推測這一變異株的N501Y會具有同樣的效果。
僅N501Y突變應該不會太影響爭對S1亞基產生的多克隆抗體,因為這些不同的抗體分子各自識別S1亞基的不同部位(
另外受到密切關注的是P681H突變。P681H在S1與S2亞基交接部位,緊鄰弗林酶(Furin)切位點PRRARS -> HRRARS。尚難判斷有多大影響。可能影響不大。
S蛋白之外,最值得一提的是發生在ORF8 上的一個轉譯終止符突變 (Q27Stop)。ORF8 是新冠編碼的病毒特異的輔助蛋白之一。新冠病毒輔助蛋白有ORF3, ORF6, ORF7和ORF8。己知ORF8對病毒複製非必需但有協助病毒適應宿主的功能。好象歐洲大陸不怎麽鬧蝙蝠和冠狀病毒。
毫無疑問各國的科學家們正在開足馬力從各個層次(
更多我的博客文章>>>