金稻米的科學依據
填補空白的技術
金稻米技術的產生基於一個簡單的 原理,水稻隻能在綠葉組織中合成 β-胡蘿卜素,而在被食用的胚乳中 卻沒有,可是這個兩步的生物合成 途徑確實存在於糧食中。僅僅通過 轉入兩種酶的基因:八氫番茄紅素 合成酶(PSY)和八氫番茄紅素脫氫酶 (crt I),糧食中可以合成β-胡蘿卜素 的途徑就能被重新激活,β-胡蘿卜 素就能因此在被食用的胚乳中製造 並積累。

類胡蘿卜素及其衍生物包括數目 龐大的分子以及大量的酶和輔助 因子。源自這一重要途徑的化合 物包括植物激素,如脫落酸(abcisic) 和赤黴素,維生素E,部分葉綠素分 子和許多天然色素。其中少量類胡 蘿卜素具有前體維生素A活性。
更詳盡的科學背景
所有能富聚類胡蘿卜素的植物組織都存 在類胡蘿卜素整合機製的體係中,包括 結晶,油沉積,膜擴散或蛋白脂質吸收 。在不合成類胡蘿卜素的水稻胚乳中由 於脂類構成比較低,因此缺乏類胡蘿卜 素沉積的條件。該技術的另一個限製是 ,在金稻米中缺乏類胡蘿卜素前體物的 供應。此外,還有許多人認為,在胚乳 中完全缺乏類胡蘿卜素生物合成途徑。
由於這些限製因素,金稻米的發展經曆 了一段滯後的階段,直至研究者積累了 足夠的科學數據,才激勵Peter Beyer和Ingo Potrykus共同 奮鬥,試圖進行這項看似無法實現的科 研努力。他們具有突破性的研究成果表 明,隻需要轉入兩個基因就能使金稻米 能在胚乳中製造胡蘿卜素,使上麵所述 及的限製條件不影響該項技術(Ye等,2000年 )。其中第一個轉基因編碼八氫番茄紅 素合成酶(PSY),通過應用內源性合成雙 香業基(geranylgeranyl)二磷酸酯,形成八氫番茄 紅素,一種帶有三烯生色基團的無色胡蘿 卜素(Burkhard等,1997年)。另一個轉基因編碼 細菌胡蘿卜素脫氫酶(CRT I)通過加入4個雙 鍵形成共軛結合。這兩個基因共同表達後 形成番茄紅素,這是一種含有十一個雙鍵 (undecaene)的生色基團的紅色化合物。在任何 不同轉化和遺傳背景的水稻中,番茄紅素 從未被觀察到,相反卻發現了α-和β-胡蘿 卜素以及大量含氧類胡蘿卜素的存在,如 葉黃素和玉米黃素。胚乳中的類胡蘿卜素 樣本顯示,兩個轉基因自身的酶活動已超 出了預期的終點。由Peter Beyer實驗室的同事撰寫 的對該機製的詳細分析在近期的植物生理 學雜誌上予以發表(Schaub等, 2005年),他們的 研究結果解釋了一些細節如下:

類胡蘿卜素生物合成的前體分子是雙香 業基二磷酸酯基。兩條實線所框出的限 速步驟因為兩個轉基因的合作表達而被 打破,因而不再需要通過去飽和酶PDS和ZDS.
一種解釋是,類胡蘿卜素途徑下遊的一些 酶,如番茄紅素環化酶(LCYs)和α -和β -胡蘿 卜素羥化酶 (hydroxylases, HYDs),可在野生型水稻胚乳 中生成,而八氫番茄紅素合成酶(PSY)以及 一種或兩種去飽和酶PDS和ZDS卻不能。轉基因作 物中由八氫番茄紅素合成酶和細菌胡蘿卜素 脫氫酶催化合成的番茄紅素的為下遊的酶提 供底物,使類胡蘿卜素的合成成為可能。事 實上,單由八氫番茄紅素合成酶的即可合成 氫番茄紅素,但不能合成去飽和酶的作用產 物(Burkhard等,1997年),這個結果為至少一個去飽 和酶PDS酶活性的缺失提供了證據。同樣,CRT I本 身並不會生產有任何顏色的水稻胚乳,因為 缺少八氫番茄紅素合成酶的(PSY)活性。
另一種解釋是,轉基因的存在反饋誘導了內 源性生物合成類胡蘿卜素的基因表達。這種 情況在隻有CRT I表達的番茄果實中得到證明。 此外,β-胡蘿卜素增加,而非番茄紅素,能 上調內源性合成類胡蘿卜素的基因表達,卻 對八氫番茄紅素合成酶(PSY)的表達產生抑製 (Römer 等,2000年)。像這種依賴於CRT I的上調機製可能 基於這樣一個事實,即作物的去飽和酶,PDS和ZDS, 共同作用產生一個四聯體形式的番茄紅素, 又稱為番茄紅素原(Bartley 等,1999年),隨後被一個 最近被確定的異構酶,番茄紅素-順式-反 式異構酶(CRTISO),異構化為反式番茄紅素 (Isaacson 等, 2002, 2004年; Park等., 2002年)。與此相反,細菌胡蘿 卜素脫氫酶(CRT I)參與作用形成特殊的全反 式番茄紅素。同樣,通過在水仙花中人工抑 製LCY, 造成全反式番茄紅素累積,從中我們能夠 持續觀察到這種特殊的類胡蘿卜素在基因和 蛋白質水平的升高,同時總的類胡蘿卜素含 量也明顯升高(Al-Babili 等, 1999年)。
含有構成表達CRT I的轉基因植物可以用來解釋金 稻米這一基因表型。通過確定內源性類胡蘿 卜素的生物合成基因的表達及產物的水平, 來獲得成功的轉基因品種。可以想象, 轉入 後表達的CRT I通過靶組織的反饋調節環路去促進 類胡蘿卜素的生物合成相關基因的轉錄激活 。這個調節環路, 由4-順式番茄紅素(tetra-cis-lycopene) 的缺失來誘發,正常來說4-順式番茄紅素是由2種 植物去飽和酶(PDS和ZDS)或者全反式番茄紅素 參與衍生而來,這個反應由CRT I催化。番茄紅素 異構體不僅代表傳統的中間體,而且還發揮 重要的調節作用。在植物和藍藻中新發現的 參與類胡蘿卜素合成途徑的酶CRTISO(Isaacson 等, 2002, 2004年; Park 等, 2002年),有可能作為調節分子對合成途徑起到 補充作用。在這種情況下,金稻米由於轉錄激 活整個類胡蘿卜素生物合成途徑而表現為橙黃 色。
在另一種情況下,有活性的下遊酶,如環化 酶和羥化酶,可能存在於野生型水稻胚乳, 這又解釋了通過轉入上遊基因,番茄紅素能 在合成後立即進入下一步合成的原因。野生 型水稻胚乳僅含低水平的葉黃素合成所需的 所有mRNA,即PSY, PDS, ZDS, CRTISO, β-LCY, ε;-LCY, β-HYD, and ε-HYD。基於PCR結果 ,同時考慮到該方法的敏感性,PSY的轉錄被有 效地抑製,這和合成途徑中,PSY表達對產生金 稻米這一基因表型,是需要但非必要的事實相 符合。 PSY單獨表達就能使金稻米中合成我們需 要的八氫番茄紅素(Burkhard等,1997年)。
熒光實時定量PCR顯示,野生型胚乳中對於CRT I的 基本要求,顯然與存在的PDS 和ZDS的轉錄相互衝突 。 這可能是由於僅表現為低水平表達的蛋白和 酶活性,而不是mRNA水平。由於低水平表達,以 及沒有放射性胡蘿卜素的底物,對PDS和ZDS複雜的 反應機製的探討是不現實的。現在的研究隻好 使用轉基因的方法進行植物體內實驗,而非體 外實驗。 胚乳特異表達的是PDS / ZDS係統,而不是CRT I ,在水稻胚乳產生可供參考比較的類胡蘿卜素 的顏色水平。 因此,水稻胚乳為植物去飽和酶 的活性提供了複雜的需求。
PDS 需要還原鏈,應用醌,一個醌還原酶,和分子 氧作為終端電子受體(Beyer等,1989年; Mayer等,1990年; Nievelstein等 ,1995年),與通過immutans突變的擬南芥識別的氧化酶 相連(見綜述, Kuntz, 2004年)。這種還原途徑對於非 綠色含類胡蘿卜素的組織尤其重要,如胚乳, 而光合電子傳遞被認為在葉綠體中發揮類似的 作用。因此,一種解釋是,當PSY作為唯一的轉基 因表達時,胚乳中由於PDS/ZDS係統表達過低而無法 產生有顏色的類胡蘿卜素。這已經表明,與PSY的 作用相比較,CRTI沒有限製反應速度,並有能力對 大量的八氫番茄紅素發揮去飽和作用,從而增 加β-胡蘿卜素的積累(Paine等,2005年)。
CRT I的主要序列與植物型的去飽和酶並無關聯。這 或許可以解釋其對輔助因子的需要,因此可能比 植物去飽和酶在胚乳中更有效的發揮作用。另一 方麵,同樣在胚乳中有活性的CRTISO,似乎也來源於CRT I (Isaacson等, 2002年; Park等., 2002年)。
PSY, PDS和ZDS在水稻胚乳中表達啟動聚順式胡蘿卜素去 飽和途徑,但β-胡蘿卜素形成主要是全反式,並 伴隨典型的順式異構體,目前,以9-順式的異構體 較多。此外,轉基因植物胚乳依靠CRT I的催化去飽 和,控製β-胡蘿卜素相同的同分異構體比值,這 表明水稻CRTISO觸發的異構體形成被控製在一個特定 的比例,與其在體外活動相符合(Isaacson等,2004年) 。CRTISO的活動對循環末端形成起了很重要的作用。 當缺乏CRTISO時,循環胡蘿卜素和衍生物葉黃素不會 在非綠葉組織中生成,這在缺失有功能的CRTISO的桔 突變番茄果實和從擬南芥CCR 2突變得到的黃化質體 中已被證明。
顯然, LCY的活性和不同類別的β-HYD的活性(見綜述 ,田和DellaPenna,2004年)依靠水稻胚乳中基因而各自表 達。目前為止所有的水稻遺傳背景的測試顯示, 對酶活動的補充在沿下遊途徑的進行中並不需要 。 此外,稻米LCYs的活動沒有被證明是限速步驟, 因為番茄紅素沒有積累。 因此,金稻米是因為內 在的大米環化酶活動呈現黃色。 熒光定量實時PCR對 轉基因水稻胚乳中分離出的RNA的分析,沒有說明在 轉基因水稻中,存在一個可能會影響類胡蘿卜素 生物合成基因所表達的反饋調節環,因為與野生 型相比,所有的相應的mRNA水平沒有改變。
盡管水稻的綠葉組織中胡蘿卜素醇含量增加,但 它與表達CRT I擬南芥和胡蘿卜素醇的構成情況基本 相似。 有這樣兩種情況,在葉黃體素下降時,β -胡 蘿卜素及其衍生胡蘿卜素醇增加,作為部分補償。 這種情況也出現在經轉化後表達CRT I的煙草BY2細胞中, 但這裏類胡蘿卜素成分沒有發生類似的變化,這可 能是因為BY2細胞不積累葉黃素和玉米黃素。在水稻 和擬南芥葉片中胡蘿卜素醇水平的變化因條件不同 有多種變化,並與CRT I表達水平呈負相關。底物向哪 一種胡蘿卜素醇形成途徑流動,是由兩個LCYs 控製的 ,LCYs參與將番茄紅素轉換成β-胡蘿卜素和α-胡蘿卜素 。 但是,用以監測類胡蘿卜素組成變化的類環化酶 的轉錄水平沒有發生重大變化。 因此,我們相信, 番茄紅素的幾何異構狀態對β- 或ε- 環的形成具有一 定影響。
缺乏CRTISO的番茄和擬南芥的突變體也會在葉片中產生 較多的β -胡蘿卜素源性胡蘿卜素醇(Isaacson等,2002年;Park 等,2002年)。因此,最有可能觀察到的是酶催化水 平和幾何異構的番茄紅素,而不是基因表達水平。 這些結果為野生型中隻存在類胡蘿卜素生物合成的 組織提供了證據,如,葉片。而在缺乏類胡蘿卜素 的野生型中的轉基因作物胚乳中則不可能觀察到。
葉片中觀察到葉黃素減少,在理論上可能降低光合 作用。作為最豐富的胡蘿卜素醇,其主要是附著在 光收集複合體II發揮作用。在擬南芥突變體中已經發 現葉黃素含量下降引起PSII天線尺寸減小(Lokstein等, 2002年 )。這表明葉黃素具有對天線結構優化的能力,同 時確保穩定有效的捕光性。因此,我們選擇在新一 代金稻米中應用組織特異性啟動子控製CRT I的表達 (Paine等 ,2005年; Al-Babili等,已投出; TTC Hoa and P Schaub, 未發表結果)。
第一代金稻米
金稻米技術第一個突破,是1999年複活期間Peter Beyer 和 Ingo Potrykus之間的合作成果(Ye 等, Science 287:303-5, 2000)。這篇論文 為在稻米中製造β -胡蘿卜素提供了證明。在 當時人們仍然相信,除八氫番茄紅素合成酶 和胡蘿卜素脫氫酶以外,還需要第三種酶--番 茄紅素環化酶去完成整個生物合成途徑。
基於已有的理論依據,科學家們立即著手發 展能提高類胡蘿卜素生產和積累的育種方法 ,因為當時人們已經認識到在相對缺乏均衡 飲食的目標人群中,合理食用量水平的金稻 米還不能完全提供每日所需的前體維生素A (1.6 微 克/克)。盡管一些東南亞地區的居民可以獲得 均衡的飲食,但很多的窮困的人卻沒有這個 條件,在實際生活中,一些邊遠地區的居民 是依靠食用大米,提供每日80%的熱量需求。
共同的努力促成後來我們所稱的第一代金稻 米的發展,也被稱為SGR1。這一代金稻米僅含 有來源於水仙花八氫番茄紅素合成酶 (PSY) 基因 和來源於噬夏孢歐文氏菌的細菌的胡蘿卜素 脫氫酶( crtI) 基因。此外,這兩個轉基因隻在水 稻胚乳表達。溫室中生長的轉基因作物產生 的類胡蘿卜素水平與預實驗結果(1-2 微克/克) 相 近,而在土地中生長的金稻米,其類胡蘿卜 素水平平均可達到6 微克/克,這可能跟作物生 長條件的改善和選種過程有關。配合每日攝 入適量的蔬菜和魚類或其他肉類(正如目前 他們的飲食攝入量),這種水平的類胡蘿卜 素含量預計將能夠滿足兒童每日推薦攝入量 。
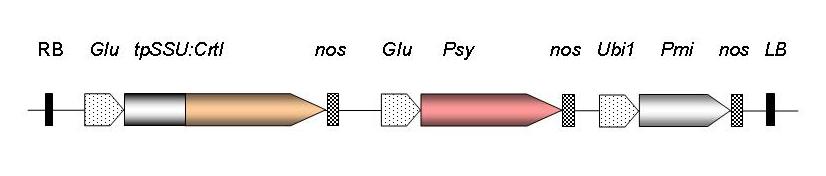
生物合成金稻米的基因結構 。RB, T-DNA 右邊界序列;穀氨酸, 水 稻胚乳特異性穀蛋白啟動子; tpSSU,豌 豆核酮糖二磷酸羧化酶的,促葉綠體 本體化小亞基轉化肽; 胭脂堿合成終止 酶; Psy, 八氫番茄紅素合成酶基因,來自 黃水仙 (GR1)或玉米(GR2); Ubi1, 玉米泛素啟動子;Pmi,磷酸甘露糖異構 酶基因來自大腸杆菌 為正選擇(GR2); LB ,T - DNA左邊界序列。
新一代金稻米
第一代金稻米,提供了為預防維生素A缺乏 症的寶貴的理念,但研究者也認識到,如 果要更有效的預防維生素A缺乏症,稻米還 需要含有更高水平的β-胡蘿卜素含量。因 為在整個過程中隻需要兩個轉基因,因此 合乎邏輯的提高金稻米中類胡蘿卜素積累 的做法是確定生物合成途徑的關鍵問題, 以及對八氫番茄紅素合成酶(PSY)和胡蘿卜 素脫氫酶(CRT I)兩種基因產物酶活性的調節 問題。 這一切可以通過應用從其他來源的同 源基因替代法或修改其調節區域去進行研究 。
在大多數多步生物合成途徑中存在一個限製 性步驟。金稻米技術的瓶頸問題就是如何調 控八氫番茄紅素合成酶(PSY)的酶活性。在 嚐試轉入來自不同來源的八氫番茄紅素合成 酶基因(PSY)後,研究者發現來自玉米和水稻 的基因能獲得最好的效果(Paine等, 2005年)。在 新一代金稻米中類胡蘿卜素的積累可多達37微 克/克,其中β-胡蘿卜素達到了31微克/克(相對 於隻有1.6微克/克 類胡蘿卜素的第一代的金稻米 有了很大的提高)。
1-3歲兒童維生素A的每日推薦量為300微克(通常 半數的每日推薦量已經足夠將維生素A維持在 正常水平)。基於β-胡蘿卜素與視黃醇當量 的比率為12 :1,隻要72克新一代的金稻米即可 提供半數維生素A的每日推薦量。根據一些國 家的稻米消耗水平,每個兒童每日進食100-200克 大米,這些金稻米提供兒童半數維生素A的每 日推薦量是完全合適的。

該圖像清楚地表明,金稻米從概念證 明階段以來,所取得的進展。新一代 的金稻米,也稱為GR2, 所含的β-胡蘿卜素 水平將能提供東南亞的健康兒童所需的 前體維生素A。