隴山隴西郡
寧靜純我心 感得事物人 寫樸實清新. 閑書閑話養閑心,閑筆閑寫記閑人;人生無虞懂珍惜,以沫相濡字字真。



博文
關於生命科學若幹數學問題的研究報告
|| 推薦到群組
關於生命科學若幹數學問題的研究報告
--對生命的本質及其發展規律的生物計量學分析
沈 律
摘 要: 隨著現代生命科學的發展,人們已經清楚地認識到生物是運動、變化和發展的。生物的運動、變化和發展是一個統一的生命過程。也可以說生物的遺傳、進化與發育是一個統一的生命運動過程。現在,隨著人們對人體生物基因組及其生物遺傳信息的深入研究,發現人體生物基因組就是一部寫了幾十億年的“生命之書”或繪了幾十億年的“生命藍圖”。而生物的遺傳、進化與發育過程就是這部“生命之書”或“生命藍圖”的不斷寫作、不斷繪製與不斷修改過程,同時也是這個“生命之書”或“生命藍圖”的不斷解讀、不斷實現與不斷選擇過程。本文首先對生物新陳代謝一般均衡定律進行分析,了解生物代謝同化與異化過程的相互關係,提出新陳代謝指數概念。再通過對人類基因組的形成過程進行研究,進一步論述和介紹了生物基因組的形成及其生物遺傳信息傳遞的中心法則以及經典遺傳學定律;其次通過對人類基因組遺傳信息的矩陣分析提出:任何一種生物其基因組在自然形成過程中都在不斷地存儲兩類生命遺傳信息,一類是縱向生命遺傳信息;另一類是橫向生命遺傳信息。此兩類信息的雜交重組遵循一定的分布規律。據此,提出生物進化的遺傳信息矩陣分布定律。最後,通過對人體生殖與發育過程的係統觀察及細胞(基因組)增長的動力學分析,又發現人體在生殖與發育過程中出現兩種類型細胞增長:一種是胚細胞(胚胎幹細胎)的增長(增殖與分化);另一種是體細胞(組織幹細胞)的增長(增殖與分化),這兩種幹細胞在增長(增殖與分化)過程中均表現出飽和現象,因此,我們認為人體生長(生殖與發育)過程遵循生命周期雙S曲線規律。通過對人類基因組生命遺傳信息的複製與表達過程進行係統研究,發現人體在胚胎幹細胞增長(基因組複製)時期主要進行的是非特異性縱向生命遺傳信息(種係遺傳信息)的表達;而在組織幹細胞增長(基因組複製)時期則主要進行的是特異性的橫向生命遺傳信息(個體遺傳信息)的表達。這兩種類型生命遺傳信息的複製與表達構成了人體生命周期的全過程。據此,我們認為加強對人類基因組的形成及其複製與表達過程的係統研究,對我們今後更好地揭示生命的本質及其發展規律具有一定的現實意義。
關鍵詞:人類基因組;胚胎幹細胞;組織幹細胞;縱向遺傳信息;橫向遺傳信息;一般均衡定律;矩陣分布定律;雙s曲線規律
The Research Report on Several key Issues of Modern Life Scienc-Thebiometrics analysis ofthe nature of life and its the the double S curve law of life cycle
SHEN LU
Abstract: With thedevelopment of modern life sciences, it has been clearly recognized thecreature is movement, change and development, biological movement, change anddevelopment is a unified life processes. you can also say that the biologicalgenetic, evolutionary, developmental life is a unified movement. now, as peoplein-depth study of the human genome and its genetic information found in thehuman genome is a biological wrote billions of years "book of life"or painted billions of years of "life blueprint." and geneticbiology, evolution and development process is this "book of life" or"life blueprint" constantly writing, constantly drawing andcontinuously changing process, but also the "book of life" or"life blueprint" constantly reading, continue to achieve and continuethe selection process. firstly, by making the formation process of the humangenome research, further discussion and presentation of the formation and thecentral dogma of genetic information transfer biological genome and mendeliangenetics;followed by the human genome genetic quasibearing matrix analysispresented: any an organism whose genome is constantly stored in two types ofnaturally formed during the life of the genetic information, one is alongitudinal genetic information of life; another kind of life is a lateralgenetic information. this reorganization of hybrid follow certain types ofinformation distribution. accordingly, the proposed genetic information matrixdistribution law of biological evolution.finally, through the humanreproductive system and developmental processes and cell growth observedkinetic analysis also found that the existence of two types of human cells togrow in the reproductive and developmental processes: one is the embryonic stemcells of fetal growth (proliferation and differentiation) ; the other is thegrowth of tissue stem cells (proliferation and differentiation), these two stemcell growth (proliferation and differentiation) process exhibited saturation,therefore, we believe that human reproductive and developmental processesfollow the laws of the life cycle of the double s curve . systematic researchthrough replication and expression processes of the human genome life ofgenetic information found nonspecific longitudinal life genetic information ofhuman embryonic stem cell growth (genome replication) time mainly (the germlinegenetic information) expression; in tissue stem cell growth (genomereplication) period is mainly carried genetic information specific horizontallife (individual genetic information) expression. these two types ofreplication and expression of genetic information of life constitutes the wholeprocess of the human life cycle. accordingly, we believe that strengthening thesystematic study of the formation and the process of replication and expressionof the human genome, to better reveal the nature of our future and thedevelopment of the law of life has a certain practical significance.
Keywords: The humangenome.embryonic stem cells,tissue stem cells,genetic informationvertical;horizontal genetic information,matrix distribution law,the law ofdouble s curve
1 引 言
生物是不斷地進化發展的,並且總是從低級到高級,從簡單到複雜。在進化發展的過程中又總是要從舊生物中不斷地產生新生物。生物的生命現象主要表現在以下幾個方麵:
第一,生物係統的新陳代謝現象。我們知道生物係統的變化發展是離不開物質、能量和信息的交換的。生物係統總是要不斷地吐故納新,新陳代謝,不斷地進行同化作用與異化作用。新生物總是要不斷地取代舊生物,在進行新陳代謝的同時還要進行生物係統的自我控製和自我調節,從而實現自身的係統目的和維持自身的相對獨立、相對穩定和相對平衡。這些構成了生物係統新陳代謝的全過程。
第二,生物係統的遺傳與變異現象。生物係統的遺傳性與變異性表現在新生物與舊生物之間相似性上,新生物在很多方麵保存舊生物的一些特征。但又不是原生物的翻版。
第三,生物係統的進化與分化現象。生物是進化發展的,生物的進化也具有一定的規律。這個規律表現在從低級到高級,從簡單到複雜,從無序到有序。
第四,生物係統的生殖與發育現象。生物的生殖過程與生物的發育過程是密不可分的。生物的生殖過程是從受精過程開始的,首先形成受孵,然後通過一個孕育過程,再通過生育的分娩的方式產生新的生命,最後發育成一個新的個體。這是高級一點生物所表現的生命現象,低級生物像細菌由於其基因組既不沒有縱向生物遺傳基因信息,也沒有橫向生物遺傳基因信息,所以它沒有高級生物的生殖與發育過程。
以上是生物係統具有的一些重要生命現象。對這些生命現象的深入研究將有助我們更好地認識生物係統和改造生物係統。
2生物新陳代謝的一般均衡定律
2.1生物新陳代謝的一般均衡現象
機體與機體內環境之間的物質、能量和信息交換以及生物體內物質、能量和信息的自我更新過程叫做新陳代謝。新陳代謝包括合成代謝(同化作用)和分解代謝(異化作用)。生物在新陳代謝過程中一方麵表現出同化現象,另一方麵表現出異化現象。同化現象表現為合成物質、吸收能量和信息的過程,而異化現象則表現為分解物質、釋放能量和信息的過程。這兩種現象促成了生物生命代謝的全部過程。

圖1 生物新陳代謝基本過程示意圖
同化作用與異化作用是兩個相互對立又相互統一的過程,在這個過程之中必然存在著一種相對平衡狀態,從微觀上講,一方麵簡單分子吸收能量合成碳水化合物、脂質、蛋白質、核酸完成同化作用,另一方麵,則又分解釋放能量轉化為簡單分子,完成異化作用。這個過程在某個特定時期必然達到均衡狀態;這是一個動態變化過程,是一個從平衡狀態走向非均衡狀態然後又走向均衡狀態的過程。這個過程,從宏觀上看,隨著胎兒的形成與人體老化的形成,都在向均衡化方向發展。這是一種從非常規均衡走向另一種是常規均衡過程,反映在生物的新陳代謝上的規律就是一般均衡規律。人體就是這樣通過非常規均衡到常規均衡再到非常規均衡以至不斷進步發展。人體新陳代謝度往往與其同化和異化之間具有一定的相關性。這種相關性主要表現為同化作用越大,其代謝指數越大,異化作用越大其代謝指數越小。也就是同化率越大,其代謝度越大;異化率越大,其代謝度越小。因此,在數學坐標係裏,同化率曲線和異化率曲線之間必然存在一個交叉點,並在一定的狀態下形成一種均衡。這個均衡點即為的生物新陳代謝均衡點,這個均衡狀態即為生物新陳代謝的一般均衡狀態。反映在生物新陳代謝上的規律就可以稱之為生物新陳代謝一般均衡規律。據此,我們可以根據這個均衡點確定生物新陳代謝度。生物新陳代謝一般均衡現象是生物一般生命現象。生物一般均衡現象都可以看成是一種非常規均衡走向常規均衡再走向非常規均衡,循環往複,以至無窮。對生物新陳代謝度的研究是一個創新性工作。過去人們對生物新陳代謝的評價主要是從定性的角度進行研究,並為此已做了大量的工作。但從定量的角度評價新陳代謝還隻是剛剛開始的事情。現在人們對生物新陳代謝評價主要是從新陳代謝的定性描述上進行。對新陳代謝的評價不能隻停留在定性之上,在定量方麵也可以大做文章。並尋找更新的評價體係和評價模型,隻有這樣才能更好地揭示生物新陳代謝的本質及其變化規律。我們根據生物新陳代謝的同化率和異化率的相互關係及其形成的均衡狀態,提出生物新陳代謝的一般均衡理論。
該理論不僅把生物新陳代謝的同化率納入生物新陳謝的評價體係和評價模型,而且還把生物新陳代謝的異化率納入生物新陳謝的評價體係和評價模型,並找到它們之間的相互均衡關係。從生物計量學角度研究生物新陳代謝的一般均衡現象盡管是一項非常複雜而艱難的科研工作,但從某種意義上講,它更能全麵的把握生物新陳代謝全過程。因此,我們認為從生物新陳代謝的一般均衡現象入手,揭示生物新陳代謝的本質及其變化發展規律,將成為未來生物新陳代謝問題研究的一個非常重要的研究方向。隻有沿著這個方向認識下去,才能更準確、更全麵地把生物新陳代謝的本質及其變化發展規律。這對正確評價和確定生物新陳代謝具有一定的重要意義。
2.2評價生物新陳代謝的兩大重要指標體係
2.2.1生物新陳代謝同化及其同化率定理
我們知道,生物在新陳代謝的過程中,並就此對生物新陳代謝同化進行評估。如果通過生物新陳代謝同化率大,就說明其新陳代謝度小,反之則大。所謂生物新陳代謝同化率就是指生物新陳代謝同化速率即變化率。生物新陳代謝同化的新陳代謝程度(P)。因此,生物新陳代謝同化率的確定對評價生物新陳代謝度無疑是一個非常重要的生物計量學指標。生物新陳代謝度(P)與生物新陳代謝同化率具有非常重要的相關關係。這種關係表現為生物新陳代謝同化率越大,其新陳代謝度越大,生物新陳代謝同化率越小,其新陳代謝度越小;新陳代謝度與生物新陳代謝同化表現出正相關關係曲線。新陳代謝度與生物新陳代謝同化之間表現出來的這種普遍規律現象我們稱之為生物新陳代謝同化率定理。生物代謝同化率(A)是指單位時間(t)內生物新陳代謝合成和吸收的物質、能量和信息的數量(Q)。
生物同化率(A)(Biological assimilation rate)=新陳代謝合成和吸收物質、能量和信息的數量(Q)/單位時間(t)

圖2 生物新陳代謝度與同化率的關係示意圖
以縱軸OP表示生物新陳代謝度,橫軸OQ表示代謝的物質、能量和信息數,新陳代謝同化率曲線AA就是按照不同新陳代謝度下相對應的物質、能量和信息數而描繪出來的曲線。曲線上的各點都表示在不同新陳代謝度下的新陳代謝同化變化率。例如新陳代謝同化率曲線上的E點就是表示,當生物新陳代謝度為Pe時,物質、能量和信息數為Qe。
2.2.2生物新陳代謝異化率及其異化率定理
我們知道,為了了解生物新陳代謝度還可以從生物新陳代謝異化率上考慮。通過對生物新陳代謝異化分析,就可以了解生物新陳代謝異化率。如果生物新陳代謝異化速度越大,就說明其異化率大,如果生物新陳代謝異化少,就說明異化率小。生物新陳代謝異化率的確定,就是通過對生物新陳代謝異化率的的統計分析得出的,生物新陳代謝異化程度。所謂生物新陳代謝異化率是指生物在新陳代謝過程中某個時期異化變化速率。生物新陳代謝異化率。生物新陳代謝異化率在某種意義上講代表了生物的新陳代謝程度,也反映了生物新陳代謝度的大小。因此,其無疑也是確定生物新陳代謝度的一個非常重要的科學計量學指標。新陳代謝度與生物新陳代謝異化率具有一定的相關性,這種相關性表現為生物新陳代謝異化率越大,其生物新陳代謝度越小,其生物新陳代謝異化率越低,其新陳代謝度越大。生物新陳代謝度與生物新陳代謝異化率之間表現出負相關性,如下曲線。生物新陳代謝度與生物新陳代謝異化率之間表現出來的這種普遍規律現象我們稱之為生物新陳代謝異化率定理。
生物異化率(D)(Biologicaldissimilation rate)=新陳代謝分解和釋放物質、能量和信息的數量(Q)/單位時間(t)
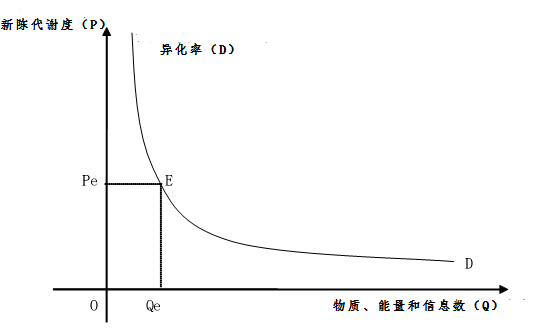
圖3 生物新陳代謝度與異化率的關係示意圖
縱軸OP表示生物新陳代謝度,橫軸OQ表示物質、能量和信息數。異化率曲線DD就是根據在每一新陳代謝度下相應的物質、能量和信息數而描繪出來的曲線。曲線上的各點表示在這一新陳代謝度上的物質、能量和信息的數量。例如,異化率曲線上的E點就是表示,當新陳代謝度為Pe時,物質、能量和信息的數量為Qe。
2.3生物新陳代謝均衡的形成
2.3.1生物新陳代謝均衡概念
生物新陳代謝均衡是一種生物新陳代謝同化率與異化率之間相一致時的生物新陳代謝度。用圖形來表示就是這同化率曲線與異化率曲線相交之點的生物新陳代謝度(圖3)。在圖中,縱軸OP表示生物新陳代謝度,橫軸OQ表示物質、能量和信息的數量。曲線DD代表論文異化率曲線,曲線SS代表同化率曲線。同化率曲線與異化率曲線的相交點E就是均衡點。E點所確定的生物新陳代謝度Pe就是該生物的均衡新陳代謝度。而E點所確定的物質、能量和信息數量Qe就是該生物達到均衡狀態的均衡物質、能量和信息數。因此,E這個均衡點既確定了均衡生物新陳代謝度,也確定了均衡物質、能量和信息數。
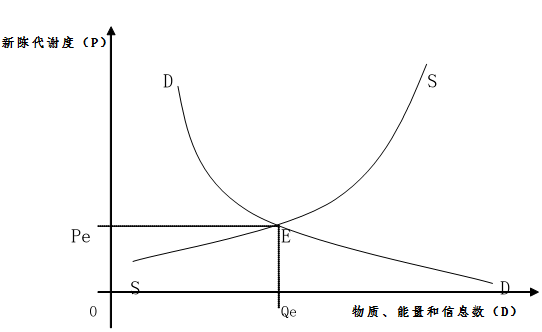
圖4 生物新陳代謝的一般均衡曲線
2.3.2 科技創新度均衡的形成
生物新陳代謝的一般均衡是生物新陳代謝同化率曲線和異化率曲線的相互作用及生物新陳代謝本身的波動而形成的。這可以從以下兩方麵來理解。
一方麵,新陳代謝度過高的情形。如果新陳代謝度過高,其同化率就會減少,而其異化率則會增加。物質、能量和信息異化數量就會多於物質、能量和信息的同化數量。出現同化率大於異化率。這樣新陳代謝就會越來越增強。隨著後繼者的不斷同化,後繼者的同類研究將會增加,隨之同化率就會不斷上升,這樣就必然導致新陳代謝度下降,最後降至一個新的均衡狀態。
另一方麵,新陳代謝度過低的情形。當新陳代謝度過低時,其新陳代謝同化率就會增加,而其異化率則會減少。物質、能量和信息的同化數量就會少於異化數量,出現異化率大於同化率。這樣該新陳代謝就會出現新的平衡。從而促進其新陳代謝度不斷上升,最後升至一個新的均衡狀態。
從以上兩種情況的分析,無論是新陳代謝度過高的情況,還是新陳代謝度過低落情況,都有動力導致新陳代謝度趨向新的均衡。因此,新陳代謝度的一般均衡過程的形成是具有其內在必然規律的,這種內在必然規律我們稱之為新陳代謝一般均衡定律。
Pd(Q)=Pa(Q)
圖3所示,縱軸OP表示新陳代謝度的大小,橫軸OQ表示異化物質、能量和信息數的多少。DD'和AA'分別是異化率曲線和同化率曲線。

圖5 生物新陳代謝均衡的動態變化示意圖
第一種情形是:當新陳代謝度為P1時,物質的吸收同化數為Q1,而物質的異化數為Q2,形成同化率大於異化率。即OQ1>OQ2。因此,從均衡理論上講,這必然導致新陳代謝度下降的壓力增大。(如由P1下降到Pe)促使同化率數量和異化率數量趨向均衡以及其新陳代謝度趨向均衡點E。形成均衡新陳代謝度Pe和均衡物質、能量和信息數量Qe。
第二種情形是:當新陳代謝度為P2時,物質的異化數為Q3,而物質的同化數為Q4,形成異化率大於同化率。即OQ3>OQ4。因此,從均衡理論上講,這必然導致新陳代謝度上升的壓力增大。(如由P2上升到Pe)促使同化率數量和異化率數量趨向均衡以及其新陳代謝度趨向均衡點E;形成均衡新陳代謝度Pe和均衡物質、能量和信息數量Qe。
根據以上生物新陳代謝的一般均衡曲線我們可以看出以下幾種情況:
第一、從一般均衡曲線上我們可以看到,E點以上部位的物質、能量和信息,由於其異化率小、同化率大,因此,其新陳代謝就強,生物價值就大。而在E點的以下部位,由於其異化率大、同化率小,因此,其新陳代謝性就弱,生物價值就小。
第二、當某物質、能量和信息異化率為零時,其同化率即反映出其新陳代謝程度;當某物質、能量和信息同化率為零時,其某物質、能量和信息異化率即反映出其新陳代謝程度。
第三、當某物質、能量和信息異化率和同化率都為零時,表明此新陳代謝度為零。
第四、衡量生物新陳代謝度的大小既取決於其異化率大小,也取決於其同化率大小。異化率和同化率可以看成是生物新陳代謝非常重要的兩種基本力量它們的相互作用決定了新陳代謝的大小。因此,新陳代謝度不僅自動調節著生物的方向,同時也促進著生物資源的不斷優化配置。
總之,在我們看來,新陳代謝均衡的形成具有其本身的內在動力機製。這種內在動力機製是生物新陳代謝本身發展規律的反映。生物新陳代謝均衡現象的發生與發展無時不受新陳代謝發展規律的控製和調節。根據異化率曲線和同化率曲線相互作用關係,我們發現,生物新陳代謝程度與其生物異化率和生物同化率具有確定性的相關性。這種相關性表現在異化率越大,其新陳代謝度越小,同化率越小,新陳代謝度越大;反之同樣。異化率和同化率曲線之間也同樣存在確定性的交叉聯係現象,並在彼此之間的交叉點上形成一種均衡,我們把這種均衡稱之為生物新陳代謝的一般均衡。而將這一均衡理論稱之為新陳代謝的一般均衡理論。
2.3.3生物新陳代謝均衡對生物自身代謝資源配置的調節
生物新陳代謝均衡的形成過程表明,生物新陳代謝均衡是生物發展的必然趨勢和正常的發展狀態。而脫離均衡點的生物新陳代謝必然導致生物新陳代謝過強或過弱、過熱或過冷的失衡狀態。通過生物新陳代謝度的形成,促進生物新陳代謝不斷地由失衡狀態轉向均衡狀態。形成生物新陳代謝的均衡協調進步的機製。這就是生物發展過程中的生物新陳代謝均衡機製。通過這種均衡生物新陳代謝狀態的形成過程,生物競爭的機製對生物代謝資源的配置發揮著積極有效的調節作用。
首先,生物新陳代謝均衡的形成是異化率與同化率波動的結果,同時也是生物新陳代謝給予兩者雙方的控製信號,是用來調節異化率與同化率之間相互關係的杠杆。當生物新陳代謝處於失衡狀態時,表明生物體係出現了生物新陳代謝過強或過弱的狀況,這樣就會對生物同化者或異化者的行為產生影響。使他們對生物新陳代謝過程中產生的調控信號做出相應的反應。並調整各自的行為,進而促使異化數和同化數以及生物新陳代謝度向均衡狀態轉變。
其次,生物新陳代謝過強或過弱,過熱或過冷都意味著生物資源配置的不合理。存在著異化率和同化率過度現象。造成生物代謝資源的浪費或利用不足。通過生物新陳代謝均衡的形成過程,生物新陳代謝機製對於代謝資源的配置不合理的狀態可以發揮有效的調節作用。使生物的發展趨勢向生物新陳代謝的均衡狀態轉化,實現生物代謝資源的合理配置。
同化率和異化率是生物新陳代謝過程中兩大基本力量,它們的相互作用決定了生物新陳代謝均衡的形成。而生物新陳代謝均衡則是生物新陳代謝的基本調節信號和杠杆,調節著同化率和異化率的平衡,促進著生物資源的優化配置的實現。同化率和異化率以及生物新陳代謝度的變化相互依存,相互影響,構成了生物進化發展過程中的核心機製——生物新陳代謝調節機製,這一機製貫穿於整個生物進化發展的始終。這一機製就好象一個無形之手,控製和調節著生物新陳代謝的整個過程。但這個無形之手也不是萬能的,醫學與防疫有形之手也將對其進行適當補充和調節。就象經濟市場規律一樣,供給與需求曲線均衡不是萬能的,這個無形之手調控著市場,但也要受到政府之有形之手的幹預和控製。
綜上所述,我們認為生物新陳代謝度與其異化率和同化率具有一定的相關性。這種相關性主要表現為異化率越大,生物新陳代謝度越小,異化率越小,生物新陳代謝度越大;同化率越大,生物新陳代謝度越大,同化率越小,其生物新陳代謝度越小。因此,異化率曲線和同化率曲線之間在坐標係裏必然存在一個交叉點,並形成一定的均衡狀態。這個均衡點即為生物新陳代謝的均衡點,這個均衡狀態即為生物新陳代謝的一般均衡狀態。據此,從生物計量學上,我們可以根據這個均衡點和均衡狀態評價生物新陳代謝的程度。生物新陳代謝均衡的形成對我們更好地揭示生物新陳代謝的本質及其發展規律具有十分重要的意義。生物新陳代謝問題是一個非常重要的學術問題。近年來引起了很多學者的高度關注,但大部分的研究主要還是從定性方麵進行描述,而進行定量分析則是近十多年來隨著生物計量學、生物數學的興起而出現。
2.4生物新陳代謝評價指標體係的構建
2.4.1生物新陳代謝度定律
根據生物新陳代謝的一般均衡規律,生物新陳代謝度與其異化率和同化率之間具有一定的相關性。這種相關性主要表現為異化越大,其生物新陳代謝度越小;異化率越小,其生物新陳代謝度越大。同化率越大,其生物新陳代謝度越大;同化率越小,其生物新陳代謝度越小。根據這個規律,我們提出:生物新陳代謝度定律(The law of biological metabolism)。就是:“生物新陳代謝度(P)與同化率(A)成正比;與異化率(D)成反比”。用數學公式表述如下:
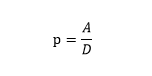
這個定律是“生物新陳代謝一般均衡規律”的數學化表達,它對我們充分認識生物新陳代謝創新一般均衡具有一定的重要作用和意義。根據這個定律,我們還可以分析生物新陳代謝度的大小,建立生物新陳代謝指數(Metabolic index,MI)生物評價指標體係。
2.4.2生物新陳代謝指數
2.4.2.2生物新陳代謝指數及其計算公式
生物新陳代謝指數(Metabolicindex,MI),又叫代謝指數。什麽是生物新陳代謝指數(Metabolicindex,MI)?所謂生物新陳代謝指數,簡單說就是指生物新陳代謝過程中,生物新陳代謝度的大小與同化率成正比;與異化率成反比。這樣就形成了一個比值,這個比值再乘一個100%所形成的一個數值就是生物新陳代謝指數。用數學公式表述如下, Metabolic index(MI)表示新陳代謝指數:
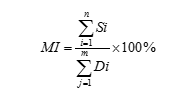

說明:即首先確定時間,然後將這個基點時間所有的同化代謝物質、能量和信息數相加,形成這個基點時間數總和。

說明:即首先確定時間,然後將這個基點時間所有的異化代謝物質、能量和信息數相加,形成這個基點時間數總和。
新陳代謝指數(MI)對新陳代謝度是相對準確的數值。這個指數就是將同化率指標體係與異化率指標體係進行均衡處理,形成一個均衡指標體係。但這個指數數據不是一成不變的,他可以形成波動。正是這種波動性,表明其動態性,持續性,發展性。
總之,我們認為,同化率和異化他率是新陳代謝過程中兩大基本力量,它們的相互作用決定了新陳代謝均衡的形成。而新陳代謝均衡則是新陳代謝的基本調節信號和杠杆,調節著同化率和異化率的平衡,促進著生物自身資源的優化配置的實現。同化率和異化率以及新陳代謝度的變化相互依存,相互影響,促使新陳代謝過程從均衡走向非均衡再走向新的均衡。從而構成了生物進化發展過程中的核心調控機製,即新陳代謝均衡調控機製,這一調控機製則貫穿於整個生物進化發展的始終。這一調控機製的揭示對生物研究事業的發展,特別是對新陳代謝的評價以及生物自身資源的合理配置具有非常重要的指導意義。這個調節機製好似“無形之手”,不斷地控製和調節著新陳代謝的進行,但這個無形之手也不是萬能的,它也將受到來自醫學和防疫“有形之手”的幹預和調節。
綜上所述,作者認為新陳代謝度與其異化率和同化率之間具有一定的相關性。這種相關性主要表現為異化率越大,其新陳代謝度越小,異化性越小,其新陳代謝度越大;同化率越大,其新陳代謝度越大,同化率越小,其新陳代謝度越小。因此,異化率曲線和同化率曲線之間在數學坐標係裏必然存在一個交叉點,並形成一定的均衡狀態。這個交叉點即為新陳代謝的均衡點,這個均衡狀態即為新陳代謝的一般均衡狀態。據此,從生物計量學上,我們可以根據這個均衡點和均衡狀態評價新陳代謝的程度。新陳代謝度評價問題不僅是一個非常重要的學術理論問題,而且也是一個非常重要的實踐問題。
3 生物基因組的形成及其生物遺傳信息傳遞的中心法則
3.1基因的分子生物學基礎
從1953年華森與克裏克發現DNA雙螺旋結構在《NATUER》發表時起,生命科學就進入了分子生物學時代。從那時起人們對生命本質的認識又進入了一個新階段。也就是把人類對生命的本質認識推進到分子水平,也使人們從分子水平認識並揭示生命的發展規律問題成為可能。現在我們知道現代生命科學就是通過以分子生物學(分子遺傳學)為主的研究生命的本質及其生命的發育規律和生命活動規律以及各種生物之間和生物與環境之間相互關係的科學。最終能夠達到認識生命,改造生命,最終駕馭生命的過程。並用這些知識為人類實現治療診斷疾病(遺傳病)、提高農作物產量、改善人類生活和保護環境等目的。

圖6 1953年華森和克裏克在《NATURE》雜誌發表DNA結構論文
說明:該論文的發表標誌著分子生物學的誕生。人們可以從分子水平上揭示生命的本質及其進化發育規律,並根據這些規律解決一係列生命科學的理論與實踐問題。
3.2 生物基因組和人類基因組計劃
現代遺傳學家認為,基因是DNA(脫氧核糖核酸)分子上具有遺傳效應的特定核苷酸序列的總稱,是具有遺傳效應的DNA分子片段。基因位於染色體上,並在染色體上呈線性排列。基因不僅可以通過複製把遺傳信息傳遞給下一代,還可以使遺傳信息得到表達。不同人種之間頭發、膚色、鼻子等不同,是基因差異所致。人類隻有一個基因組,大約有2-3萬個基因。每個人的基因組都是不相同的。因此,表現出差異性。
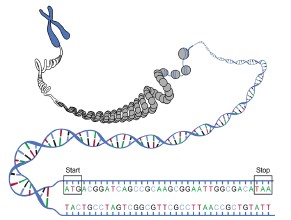
圖7 生物基因組結構示意圖
人類基因組計劃是美國科學家華森於1985年率先提出的,旨在人類基因組闡明30億個堿基對的序列,發現所有人類基因並搞清其在染色體上的位置,破譯人類全部遺傳信息,使人類第一次在分子水平上全麵地認識自我。計劃於1990年正式啟動,這一價值30億美元的計的目標是,為30億個堿基對構成的人類基因組精確測序,從而最終弄清楚每種基因製造的蛋白質及其作用。

圖8 國際人類基因組計劃協調人柯林斯
弗朗西斯·柯林斯(Francis Collins Ph.D., M. Phil & M.D.),醫學博士、遺傳學家。美國國立衛生院(NIH)國家人類基因組研究中心主任,美國醫學院院長,國家科學院院士。由他帶領的工作小組曾在發現囊性纖維症、神經纖維瘤和Huntington舞蹈症,及一種特殊類型成人白血病的基因的研究中做出重大的貢獻。1993年柯林斯博士成為美國國家人類基因組中心的第二任負責人,成為詹姆斯沃森的繼任者,並擔任國際人類基因組計劃協調人。
打個比方,這一過程就好像以步行的方式畫出從北京到上海的路線圖,並標明沿途的每一座山峰與山穀。雖然很慢,但非常精確。隨著人類基因組逐漸被破譯,一張生命之圖將被繪就,人們的生活也將發生巨大變化。基因藥物已經走進人們的生活,利用基因治療更多的疾病不再是一個奢望。因為隨著我們對人類本身的了解邁上新的台階,很多疾病的病因將被揭開,藥物就會設計得更好些,治療方案就能“對因下藥”,生活起居、飲食習慣有可能根據基因情況進行調整,人類的整體健康狀況將會提高,二十一世紀的醫學基礎將由此奠定。利用基因,人們可以改良果蔬品種,提高農作物的品質,更多的轉基因植物和動物、食品將問世,人類可能在新世紀裏培育出超級物作。通過控製人體的生化特性,人類將能夠恢複或修複人體細胞和器官的功能,甚至改變人類的進化過程。人類基因組計劃的目的。測出人類基因組DNA的30億個堿基對的序列,發現所有的人類基因,找出它們在染色體上的位置,破譯人類全部遺傳信息。這項計劃已於2001年完成基因組草圖繪製任務。
3.3 生物基因組的形成過程
隨著現代生命科學的發展,人們已經清楚地認識到生物是運動、變化和發展的。生物的遺傳、進化、發育過程具有一定的相關統一性。現在,隨著人們對人體生物基因組及其生物遺傳信息的深入研究,發現人體生物基因組就是一部寫了幾十億年的“生命之書”或繪了幾十億年的“生命藍圖”。而生物進化與生物發育過程就是這部“生命之書”或“生命藍圖”的不斷寫作、不斷繪製與不斷修改過程,同時也是這個“生命之書”或“生命藍圖”的不斷解讀、不斷實現與不斷選擇過程。現代生物科學發現,一個受精卵之所以能夠按照嚴格的順序發育成生物個體,其根據就是受精卵中有一個完整的基因組圖譜即細胞核中的全部遺傳基因(全部生命遺傳信息),發育的程序都已編碼於基因組中。發育程序可和一個建築物的藍圖相比擬。發育的過程是通過遺傳基因信息有秩序表達而使藍圖逐步實現的過程。那種認為精子或卵子中有一個完整的小生物,把發育過程看成是簡單的長大或展現,而否定進化發展的先成論顯然是錯誤的。但是細胞中確實存在著一種先成的因素,即發育藍圖,這點也是確定無疑的。然而,人體係統的發育則又是嚴格按照這個藍圖來實現的,是一個漸成的過程,這點同樣也是無可爭議的。這就是現代生物學理論對傳統“先成論”和“漸成論”的一種新解釋,並且這種解釋則將上述兩種理論進行了有效的統一,當然不是傳統意義上的統一。根據上述理論,我們認為,人體的生殖與發育過程可以看成是人體生物遺傳基因組的複製與表達過程。在這個過程中,首先,是建立一個新的人體生物基因組圖譜(受精卵),繪製好一個人一生的生命周期藍圖。然後,在此基礎上進行人體生物基因組的複製和表達,最後,完成人體生長發育全過程。
人類基因組圖譜在自然情況下是如何繪製的?其中又包含那些生命遺傳信息?我們認為,人體的生命遺傳信息全部存儲於基因組DNA結構序列之中,人類生物基因組圖譜在作者看來是這樣形成的。從縱向看,人類生物基因組是從生物界一步步進化而來,是生物進化的產物;但從橫向看,人類生物基因組則是親代遺傳基因即親代配子染色體雜交的產物。這一縱一橫繪成了我們今天人類基因組圖譜。所以,我們認為,人體生命遺傳信息主要來源於兩個方麵的遺傳:一方麵,人類生命遺傳信息來自於生物界的長期進化,是從整個生物界繼承而來;另一方麵,人類生命遺傳信息又來自於親代染色體的雜交,是從親代父係和母係身上繼承而來。因此,我們人體基因組圖譜中應該包含兩方麵的生命遺傳信息,即一方麵是縱向的種係進化信息即縱向生命遺傳信息(一般共同信息,即種係遺傳信息);另一方麵是橫向的父母遺傳信息即橫向生命遺傳信息(特殊差異信息,即個體遺傳信息)。不管是縱向生命遺傳信息,還是橫向的生命遺傳信息,在遺傳基因的結構序列上和存儲數量上均發生了一定質與量的變異。這點是非常至關重要的,正是因為變異現象的出現使得人類生物體可以不斷地去適應更為複雜的外界環境,同時也使人類與動物界之間,人類個體與個體之間產生生物性狀的差異。正是這種差異導致豐富多彩的生命世界的形成。所以,生物進化最本質的方麵是生物基因組的進化,生物從低級到高級,從簡單到複雜,主要區別在於生物基因組結構與功能的差異。因此,可以說生物基因組的進化是生物進化的根本所在,生物的進化與生物基因組的進化直接相關。
總之,我們認為,對生物基因組縱向生命遺傳信息(一般共同信息,即種係遺傳信息)和橫向生命遺傳信息(特殊差異信息,即個體遺傳信息)的認識及其基本概念的引入是非常重要的。它將有助於我們更全麵地,更準確地對各種生物遣傳、進化和生物發育過程進行定量分析。不同的生物因為其基因組結構序列不同,其所包含的縱向生命遺傳信息和橫向生命遺傳信息的質與量也是不一樣的。因此,其生物進化或生物發育的層次和位置也就不一樣。低等生物與高等生物相比較,之所以表現出生物進化和生物發育的差異性以及表現出生物個體結構和功能的差異性,其原因就在於此。生物進化與生物發育過程既是生物基因組內縱向生命遺傳信息與橫向生命遺傳信息的存儲與重組過程,又是這兩方麵生命遺傳信息的複製與表達過程。生物從低級到高級,從簡單到複雜,由於其生物基因組所包含的上述兩類信息的質與量的不一樣而分布於不同的進化與發育層次之中並表現出不同的分布狀態。這就是我們對生物基因組內縱向生命遺傳信息和橫向生命遺傳信息的存儲與重組、複製與表達過程所具有的生物進化和生物發生學意義的理解。
3.4 生物遺傳信息傳遞的中心法則
現代生命科學研究發現,生物細胞核DNA是存儲生命遺傳信息的主要載體。DNA通過複製與轉錄的方式把遺傳信息傳遞給RNA,RNA再通過一定的翻譯程序把生命遺傳信息傳給蛋白質,這就是生物遺傳生物遺傳信息傳遞的中心法則。現代生命科學研究進一步證實人類生物基因組所包含的各類生命信息都在通過中心法則進行傳遞。
人類的全部生命遺傳信息都存儲在基因組中人類基因組是整個生命遺傳信息的載體。現代生命科學發現人類基因組中的生命遺傳信息支配著人體發育的全部過程。生物基因組一旦形成其所存儲的生物遺傳信息就要進行傳遞。
生物的遺傳、進化和發育過程都受生物基因組遺傳信息的調控。生物基因組生物遺傳信息的複製與表達是通過一定的方式傳遞的,生物信息的傳遞過程是遵循一定的基本法則的。早在二十世紀50年代克裏克就提出:"生物信息傳遞的中心法則"。
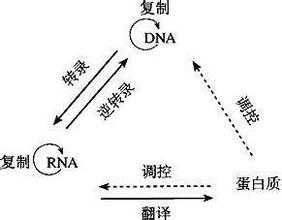
圖9 人體生物遺傳信息傳遞中心法則
4 現代生物遺傳學四大定律
各種生物都能通過生殖產生子代,子代和親代之間不論在形態構造或生理機能的特點上都很相似,這種現象就叫遺傳。但是,親代和子代之間,子代各個體之間不會完全相同,總會有所差異,這種現象叫變異。遺傳和變異是生命的重要特征。遺傳與變異導致生物發育與分化或進化。發育與分化或進化也是生物生命重要特征。
4.1 生物遺傳因子的分離定律
孟德爾(GregorJohann Mendel)1822年7月20日出生於奧地利帝國西裏西亞海因策道夫村,在布隆(Brunn)(現在是捷克的布爾諾 )的修道院擔任神父,是遺傳學的奠基人,被譽為現代遺傳學之父。他通過豌豆實驗,發現了遺傳學三大基本規律中的兩個,分別為分離規律及自由組合規律。

圖10 孟德爾(Gregor Johann Mendel,1822-1884)
孟德爾認為,生物性狀是由遺傳因子(基因)控製的,每種生物有許多性狀,因此,每種生物有許多遺傳因子(基因)。生物的一對性狀是由兩個遺傳因子(基因)---其中一個來自父本,一個來自母本---決定的。決定一對性狀的兩個遺傳因子(基因)可以獨立存在,相互分離。後來人們把孟德爾發現的這個遺傳定律稱之謂分離定律。又叫遺傳學第一定律:即一對遺傳因子(基因)在雜合狀態下保持相對獨立性,當配子形成時,可以各自分開。通過圖示可以更好地理解孟德爾的分離定律,假如高植株豌豆的高杆性狀的決定因子用D表示,相對的,矮杆性狀的決定因子用d表示,產生的子一代和子二代因子分離情況。
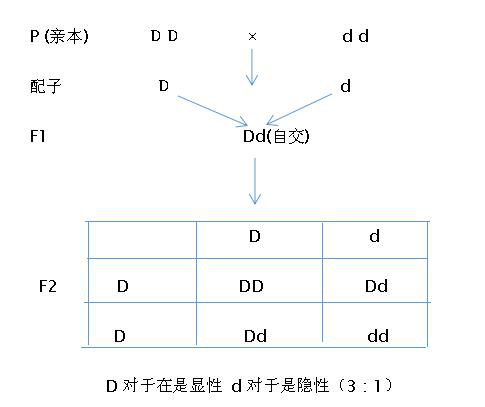
圖11 生物遺傳因子的分離定律
3.2 生物遺傳因子的自由組合定律
孟德爾在分析了一對相對性狀傳遞規律的基礎上,再利用具有兩對性狀差異性狀放在一起,發現了新的遺傳定律,即後來人們稱之謂自由組合定律。
孟德爾在雜交試驗中,親本之一是子葉黃色、飽滿;之二是子葉綠色、皺癟。得到的子一代全部是子葉黃色、飽滿。子一代自花授粉,得到子二代,共計556粒。其中有黃滿、綠皺,還有黃皺、綠滿,後兩種性狀是親代所沒有的。為什麽會出現後兩種性狀呢?這是因為黃和綠、滿和皺各是一對相對性狀,在形成配子時,它們可以相互分離,與不同對的相對性又可以相互結合。
根據性狀的分離規律,在子二代裏,應有3/4黃的和1/4綠的,而在3/4黃的裏應有3/4滿的和1/4皺的,在1/4綠的裏應有3/4滿的和1/4皺的.這樣即有:
黃滿 3/4 × 3/4=9/16
黃皺 3/4 × 1/4=3/16
綠滿 1/4 × 3/4=3/16
綠皺 1/4 × 1/4=1/16
孟德爾雜交試驗中,子二代556粒,黃皺和綠滿都是101粒,綠皺是32粒,正好接近9:3:3:1這個比數。
上述雜交結果如何解釋呢?子葉的黃和綠、滿和皺分別是一對相對性狀,設親本黃滿的遺傳型是YYRR、綠皺的遺傳型是yyrr。它們的配子分別是YR、yr,得到的子一代其遺傳型是YyRr。子一代自交時,形成雌、雄配子YyRr在形成配子時有四種可能的組合:YR、Yr、yR、yr,各種條件不變,產生的配子數應該是1:1:1:1。4種雄配子和4種雌配子結合,可有16種組合。因此在子二代中有9種遺傳型,4種表現型(如圖3所示)。孟德爾自由組合定律可表達為:兩對基因在形成配子時,是彼此分離,自由組合的。

圖12 生物遺傳因子的自由組合定律
4.3 生物遺傳基因的連鎖交換定律
托馬斯·亨特·摩爾根 (Thomas Hunt Morgan) 是美國進化生物學家,遺傳學家和胚胎學家。發現了染色體的遺傳機製,創立染色體遺傳理論, 是現代實驗生物學奠基人。1866年9月25日出生在肯塔基州的列克星敦(Lexington)。在肯塔基州立學院 (State College of Kentucky) 現在的肯塔基大學(University of Kentuck)接受教育。他在約翰霍普金斯大學(Johns Hopkins University)研究胚胎學,並於1890年獲得博士學位。1945年 12月4日,因動脈破裂,摩爾根在帕薩迪納逝世,享年78歲。於1933年由於發現染色體在遺傳中的作用,贏得了諾貝爾生理學或醫學獎。托馬斯·亨特·摩爾根 (Thomas Hunt Morgan)提出了連鎖交換定律,後被人們稱之謂遺傳學第三定律。基因的連鎖與互換規律。的定義為:生殖細胞形成過程中,位於同一染色體上的基因是連鎖在一起,作為一個單位進行傳遞,稱為連鎖律。在生殖細胞形成時,一對同源染色體上的不同對等位基因之間可以發生交換,稱為交換或互換律。(law of linkage and crossing-over)

圖13 摩爾根(Thomas Hunt Morgan,1866-1945)

圖14生物遺傳基因的連鎖和交換定律
連鎖和互換是生物界的普遍現象,也是造成生物多樣性的重要原因之一。一般而言,兩對等位基因相距越遠,發生交換的機會越大,即交換率越高;反之,相距越近,交換率越低。因此,交換率可用來反映同一染色體上兩個基因之間的相對距離。以基因重組率為1%時兩個基因間的距離記作1厘摩(centimorgan,cM)。
交換值(重組率):指同源染色體的非姊妹染色單體間有關基因的染色體片段發生交換的頻率,一般利用重新組合配子數占總配子數的百分率進行估算。
交換值(%)=(重新組合配子數/總配子數)×100
4.4生物遺傳基因的矩陣分布定律
沈律(1962-)男、安徽人、中國科技大學畢業、研究生學曆、中國自然辯證法研究會科學技術學專業委員會委員;皖南醫學院科學技術學研究所所長,曾任皖南醫學院恩普科技谘詢公司經理;中國管理科學研究院研究員、中國科協《發現》雜誌社理事;研究方向:科學計量學、經濟計量學、社會計量學、科學學與科技管理;生物計量學(生物數學)、生命科學與醫學。其主要學術貢獻:第一,發現“生物新陳代謝一般均衡定律”,這是指生物新陳代謝過程中,同化作用與異化作用之間普遍存在著一般均衡關係;第二,發現“生物遺傳第四大定律”,即“生物遺傳基因矩陣分布定律”,這是指出生物基因組中存在縱向生物遺傳基因和橫向生物遺傳基因,這兩組基因雜交形成一個矩陣分布序列,這個矩陣分布序列決定了生物進化和發育(分化)的程度,決定了生物進化與發育(分化)基本規律過程。第三,發現“生命周期雙S曲線規律”,這個規律與生物遺傳基因矩陣分布規律之間有著一種必然的內在聯係機製。這種聯係製是由生物遺傳基因矩陣分布定律所決定的。這三大基本定律(規律)的發現,都是在前人工作基礎上完成的,但都是對前人工作的突破與發展,因此,它在生命科學(生物學)中與前麵生物學幾大定律(規律)發現具有同等重要的學術意義和價值。

圖15 沈律(shenlu,1962-)
4.4.1 生物(種係與個體)遺傳因子自由分離與組合規律
盡管前麵三大遺傳定律已經被人們所熟悉,但我還要在這裏提出第四大生物遺傳定律,那就是生物遺傳因子矩陣分布定律。因為它揭示了生物的種係與個體遺傳規律。生物的種係與個體遺傳因子的自由分離與自由組合定律具體地闡述了生物體係可以通過不同生物遺傳因子的相互作用、相互交叉、相互聯係、相互轉化、相互轉換、相互重組、相互連鎖,等等方式產生豐富的遺傳性與變異性。而這一切正是導致生物係統遺傳與變異現象發生的根本原因,它也是生物進化與發育(分化)的根本動因。因此,這個定律與前麵三大遺傳定律具有同等的生物學價值和意義
生物遺傳因子具有代表性的主要有二種類型,即:X(種係生物遺傳因子,又叫一般生物遺傳因子、縱向生物遺傳因子)、Y(個體生物遺傳因子,又叫特殊生物遺傳因子、橫向生物遺傳因子),但隨著生物的不斷進化,生物的結構層次就越來越複雜,越來越增加,這樣其生物遺傳因子的數量也將形成一個集合序列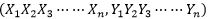 隨著生物不斷增加,使得生物遺傳因子在生物體係的內部不斷地自由分離與組合,不斷地連鎖與交換。從而導致各種各樣生物個體的形成。
隨著生物不斷增加,使得生物遺傳因子在生物體係的內部不斷地自由分離與組合,不斷地連鎖與交換。從而導致各種各樣生物個體的形成。
按照生物遺傳因子的自由分離與自由組合規律,2對生物遺傳因子 的可能組合方式為4,其組合數為16。3對生物遺傳因子
的可能組合方式為4,其組合數為16。3對生物遺傳因子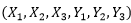 的可能組合方式為8,其組合數為64;4對生物遺傳因子
的可能組合方式為8,其組合數為64;4對生物遺傳因子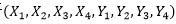 的可能組合方式為16,其組合數為256;n對生物遺傳因子
的可能組合方式為16,其組合數為256;n對生物遺傳因子 的可能組合方式為2n,其組合數為 2n*2n。由於自然界生物遺傳因子的數目是很多很多的,而且又大都為雜合型,因此,在進化過程中其生物遺傳因子的重組就會產生多種多樣,千變萬化,豐富多彩的新生物係統。
的可能組合方式為2n,其組合數為 2n*2n。由於自然界生物遺傳因子的數目是很多很多的,而且又大都為雜合型,因此,在進化過程中其生物遺傳因子的重組就會產生多種多樣,千變萬化,豐富多彩的新生物係統。
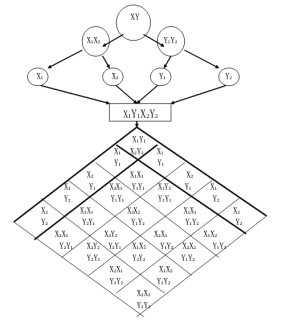
圖16 生物(種係與個體)遺傳基因自由分離與組合規律示意圖
生物之所以發生遺傳現象與變異現象,是因為生物體係為了在外界環境中適應、生存和發展就必須與外界環境進行物質、能量和信息的交換,由此形成具有一定特征的各種類型的生物所組成的生物係統。這個係統具有一定的結構與功能,並且這種結構與功能的出現意味著一種生物的形成。任何一個生物體係都具有自己的特定結構與功能,都具有自己的特定生物規範。總體生物係統有總體生物的特定生物規範,個體生物有個體生物的特定生物規範。一般生物有一般生物的特定生物規範,特殊生物有特殊生物的特定生物規範。各種生物體係的生物規範都具有本質上的區別與聯係。而生物體係的規範則又是由生物基因所決定的。所以生物遺傳因子的自由分離與組合,連鎖與交換是導致生物體係遺傳現象與變異現象發生的根本原因。由此我們不難看出,生物體係之所以發生遺傳現象,是因為生物體係在形成與發展過程中,新生物體係是從舊生物體係中分裂出來,但在其生物規範中仍然保留了舊生物規範中的生物遺傳因子,舊生物遺傳因子仍然在不同程度上參與了新生物規範的組合,這就是導致生物體係遺傳現象發生的主要原因。而變異現象的發生,則是由於在環境因素的作用下,導致生物遺傳因子無論在質上還是在量上,都發生了很大的變化。使得參與組合新生物規範的生物遺傳因子不是原來意義上的回歸重現,這樣就自然導致生物體係的變異。生物體係的遺傳現象與變異現象之所以發生,一方麵取決於生物體係內部新舊生物遺傳因子的自由分離與自由組合,另一方麵取決於生物體係外部環境的變化與選擇,是生物體係內外兩方麵因素作用的結果。生物遺傳因子自由分離與自由組織將導致大量的生物的產生,同時也將導致多種多樣,千變萬化,豐富多彩的生物群體的產生。
生物進化與生物發生過程既是其生物基因組的形成與發展過程,也是其生物基因組複製與表達過程。任何一種生物由於其基因組中所包含的縱向生命息與橫向生命信息的質與量的不同。而表現出不同的生物進化位置和生物發生的狀態。並分布於一定的生物進化或生物發生層次之中。從低級到高級,從簡單到複雜。通過研究我們發現每種生物的基因組其生物遺傳信息的矩陣分布特征是不一樣的,這種差異性決定了生物進化程度與發育程度。
4.4.2 生物基因組遺傳信息矩陣分布特征圖
通過對生物基因組所包含的縱向生命遺傳信息(一般共同信息,即種係遺傳信息)和橫向生命遺傳信息(特殊差異信息,即個體遺傳信息)的質與量的分析我們發現"生物基因組遺傳信息矩陣分布定律",現將該定律的具體內容敘述如下。
首先我們設定:
A表示生物基因組內縱向生命遺傳信息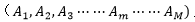 ,從低級到高級形成一組序列;
,從低級到高級形成一組序列;
B表示生物基因組內橫向生命遺傳信息 ,從簡單到複雜形成另一組序列;
,從簡單到複雜形成另一組序列;
兩類生命遺傳信息的交叉形成以下矩陣,各種生物由於其基因組中縱向生命遺傳信息(一般共同信息,即種係發育信息)和橫向生命遺傳信息(特殊差異信息,即個體發育信息)質與量的不一樣而出現不同的矩陣分布。

圖17 生物基因組遺傳信息矩陣分布特征圖
說明:根椐以上矩陣分布圖,我們認為,每種生物由於其基因組內所含的縱向生命遺傳信息和橫向生命遺傳信息的不同,而導致其矩陣分布圖的不一樣。由低級到高級,由簡單到複雜形成一定的矩陣分布序列。其數學表達式如下:

式中:S表示生物基因組總體遺傳信息矩陣分布;s表示生物基因組部分遺傳信息矩陣分布; Xij為對應點上生物基因組中遺傳信息數的統計數字;M表示總體縱向生命遺傳信息序列數,(M為正整數,M大於m);N表示總體橫向生命遺傳信息序列數,(N為正整數,N大於n)m表示部分縱向生命遺傳信息序列數;(m為正整數,m小於M)n表示部分橫向生命遺傳信息序列數。(n為正整數,n小於N)
我們認為各種生物由於其生物基因組中所包含的縱向生命遺傳信息(一般共同信息,即種係遺傳信息)和橫向生命遺傳信息(特殊差異信息,即個體遺傳信息)質與量的不同而表現出不同的矩陣分布狀態,這種分布狀態表現出從低級到高級,從簡單到複雜矩陣分布趨勢,我們把這種趨勢分布規律稱為“生物基因組遺傳信息矩陣分布定律”又稱“沈氏遺傳定律”。我們認為“生物基因組遺傳信息矩陣分布定律”是一個非常重要的生物學發現,因為它將有助於我們更好地確定各種生物進化層次和生物發育狀態。這對我們今後更準確地進行生物進化度和生物分化度的定量分析具有重要實用價值。
4.4.3 生物基因組遺傳基因(信息)特征分析
4.4.3.1 生物基因組遺傳基因(信息)矩陣特征分析
(1)通過矩陣分析,我們可以看出矩陣係統中含有兩類遺傳信息:一類是縱向遺傳信息
 ;另一類是橫向遺傳信息
;另一類是橫向遺傳信息 ;這兩類遺傳信息之和為M+N。假設兩類遺傳信息進行一次雜交形成一個生物基因組,則可從矩陣中求出這個生物基因組中理論上的交叉基因數為M×N。實際基因組矩陣中可以統計出的所有雜交基因數之和為:
;這兩類遺傳信息之和為M+N。假設兩類遺傳信息進行一次雜交形成一個生物基因組,則可從矩陣中求出這個生物基因組中理論上的交叉基因數為M×N。實際基因組矩陣中可以統計出的所有雜交基因數之和為:
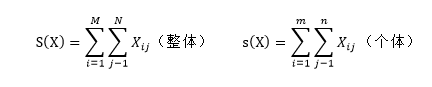
(2)每個行(橫向)、列(縱向)所代表的生物基因組內已雜交的遺傳基因數可由下式表達:


(3)兩類遺傳基因Ai與Bj間交叉出的遺傳基因數即為矩陣中對應點元素的數字:Xij

4.4.3.2 生物基因組遺傳信息量的特征分析
通過對生物基因組遺傳信息的縱向生物遺傳信息(種係遺傳信息)與橫向生物遺傳信息(個體遺傳信息)的認識,我們根據信息科學理論原理對生物基因組信息量定量分析如下:

生物基因組內縱向生命遺傳信息(種係遺傳信息)與橫向生命遺傳信息(個體遺傳信息)量之和為H(A)+H(B)。而兩類生命遺傳信息雜交後形成基因組的遺傳信息量則為H(A,B),因此,生物基因組的實際遺傳信息含量應該為:H(A,B)。
(1)縱向生命遺傳信息(種係遺傳信息)量分析,我們設定,縱向生命遺傳信息(種係遺傳信息)量為:
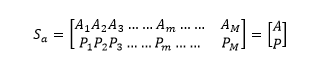
其中 代表縱向生命遺傳信息(種係遺傳信息)整體,
代表縱向生命遺傳信息(種係遺傳信息)整體,
 是縱向生命遺傳信息(種係遺傳信息)A的概率分布,Sa表示縱向生命遺傳信息結構。
是縱向生命遺傳信息(種係遺傳信息)A的概率分布,Sa表示縱向生命遺傳信息結構。
采用對數作為不定性的度量,則:
A1可能結果的不定性的量為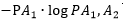 可能為
可能為 ,A3可能為
,A3可能為 一直到
一直到 ,而整個生物基因組縱向生命遺傳信息(種係遺傳信息)的不定性的量則是它們的和。其公式為:
,而整個生物基因組縱向生命遺傳信息(種係遺傳信息)的不定性的量則是它們的和。其公式為:
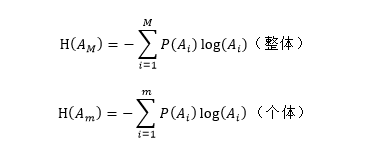
H(A)代表生物基因組縱向生命遺傳信息(種係遺傳信息)量。
(2)橫向生命遺傳信息(個體遺傳信息)量分析,我們設定,縱向生命遺傳信息(種係遺傳信息)量為:

其中 代表橫向生命遺傳信息(個體遺傳信息)整體,
代表橫向生命遺傳信息(個體遺傳信息)整體,
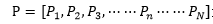 是橫向生命遺傳信息(個體遺傳信息)B的概率分布,Sb表示橫向生命遺傳信息結構。采用對數作為不定性的度量,則:
是橫向生命遺傳信息(個體遺傳信息)B的概率分布,Sb表示橫向生命遺傳信息結構。采用對數作為不定性的度量,則:
B1可能結果的不定性的量為 可能為
可能為 可能為
可能為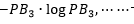 一直到
一直到 而整個生物基因組橫向生命遺傳信息(種係遺傳信息)的不定性的量則是它們的和。其公式為:
而整個生物基因組橫向生命遺傳信息(種係遺傳信息)的不定性的量則是它們的和。其公式為:

H(B)代表生物基因組橫向生命遺傳信息(個體遺傳信息)量。
4.4.4 生物進化度(分化度)比值分析
4.4.4.1物進化度(分化度)矩陣比值分析
(1)生物基因組縱向遺傳基因矩陣比值:
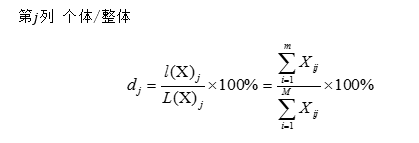
(2)生物基因組橫向遺傳基因矩陣比值:
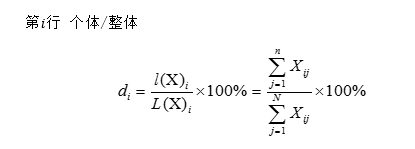
從生物進化角度上看,任何一種生物由於其基因組進化層次的不同,其基因組複製與表達的層次也就不一樣,從低級到高級,從簡單到複雜,從無序到有序。這樣在生物發育過程中也表現出不同的發育階段,出現不同的分化度。在任何一個基因組中,縱向遺傳基因Ai和橫向遺傳基因Bj雜交所形成的生物基因組的進化度(分化度),如果以Dij表征。那麽Dij即為該基因組已進化的(已分化)的基因組數(個體)與整個係統基因組總數(整體)之比:

4.4.4.2 生物基因組遺傳信息量比值分析
(1)生物基因組縱向生命遺傳信息量之比:

(2)生物基因組橫向生命遺傳信息量之比:

(3)生物基因組個體與整體生命遺傳信息量之比:

總之,我們認為,生物的進化過程從理論上講可以看成是生物基因組的進化過程,因此,對各種生物的基因組內所含有的信息量進行定量分析、比較,就可以確定某種生物的進化(分化)程度。
4.4.5 生物進化度(分化度)矩陣分析
由於Dij是矩陣D的一個元素。因此,生物進化度(分化度)矩陣D為:

生物進化度(分化度)矩陣F實際上為我們提供並展示了一幅現實的生物進化或生物發育圖景,如果將這一矩陣(D)進行模糊數學分析和計算機繪圖處理,從理論上講即可得到一幅生物進化或生物發育狀況分布圖。這樣一來,我們通過對生物進化度(分化度)的定量分析,就完全可以確定某種生物或物種的進化(分化)程度以及它們所處的生物進化層次和生物發育狀態,從而也即可確定每一種生物或物種在整個生物係統的進化位置。
4.5 現代遺傳學四大定律的區別與聯係
在進行減數分裂形成配子時,位於同一條染色體上的不同基因,常常連在一起進入配子;在減數分裂形成四分體時,位於同源染色體上的等位基因有時會隨著非姐妹染色單體的交換而發生交換,因而產生了基因的重組。應當說明的是,基因的連鎖和交換定律與基因的自由組合定律並不矛盾,它們是在不同情況下發生的遺傳規律:位於非同源染色體上的兩對(或多對)基因,是按照自由組合定律向後代傳遞的,而位於同源染色體上的兩對(或多對)基因,則是按照連鎖和交換定律向後代傳遞的。生物基因矩陣分布定律與自由分離和組合定律就更不矛盾了,它是多基因生物體基因的排列與組合規律。它反映了生物遺傳基因如何控製生物發育和進化過程。染色體內存在縱向生物遺傳基因和橫向生物遺傳基因,在基因表達過程中,首先是以縱向生物遺傳基因表達為主,因此在胚胎時期出現海克爾重演現象,出現一般生物特征,沒有特異性;出身後,以橫向生物遺傳基因表達為主,出現個體特征性狀。有特異性;基因組如果進化的比較低級,那麽就不會出現上述現象。第四大定律揭示了生物進化與生物發育的基本原因。從孟德爾遺傳定律(生物遺傳基因自由分離與自由組合定律)到摩爾根遺傳定律(生物遺傳基因連鎖和互換定律)再到沈律遺傳定律(生物遺傳基因矩陣分布定律),這些規律都是一脈相承的,它們都是用排列和組合的方式一個比一個更全麵深入地了解生物遺傳基因在個體和群體的分布情況。從而揭示了生命遺傳現象的本質及其進化發展規律。
5 人體生命周期雙S曲線規律
人體作為一個生命體係,具有生命現象,人體是生物界進化發展的最高形式。人體生殖與發育過程就是整個生物係統進化發展過程短暫而迅速的重演。人體作為一個生物個體,從其受精卵形成時起,就開始了它的生命曆程。我們把人體從受精卵形成開始,到胚胎孕育,再到生長發育,最後衰老消亡所完成的生命曆程稱為人體的生命周期。人體的生命周期過程主要包括兩個階段:第一是人體的生殖分化階段;第二是人體的生長發育階段。其生殖分化階段表現的特點是在母親生命體係內由受精、孕育,然後一步一步地形成新的生命幼體---胎兒,即完成由“受精卵---胚胎發育---胎兒形成”;其生長發育階段表現的特點是新的生命體係---新生兒,不斷地由幼稚到成熟的發育和生長,直到最後衰老消亡。即完成由“新生兒---生長發育---衰老消亡”。這兩個過程的結合正好構成人體由產生到發展再到消亡的全部曆程。這個過程表現為人體作為一個生命係統發展的一次完整的“生命周期”。
5.1 人體生殖與發育的基本過程
人體的整個生命周期過程就是人體的生殖與發育過程。我們把這個過程分為兩個階段,八個時期。即:一、生殖分化階段,其中包括:A、受精卵形成時期,B、受精卵卵裂時期,C、三胚層形成時期,D、胎兒形成時期;二、生長發育階段,其中包括:E、新生兒誕生時期,F、體細胞增殖時期,G、性成熟時期,H、人體衰老死亡時期。現分述如下:
5.1.1 生殖分化階段
A、 受精卵形成時期:此期是胚細胞(基因組)增長的延滯期。人體兩個性細胞,即配子(精子、卵子)融合為一,成為合子或稱受精卵。而配子(精子與卵子)是由母細胞經減數分裂而產生的。在減數分裂過程中,配子母細胞分裂兩次,但DNA隻複製一次,由於少了一次DNA複製,因而兩次分裂產生的4個細胞(配子)各隻含有半數的染色體。例如人的體細胞含有23對染色體,減數分裂後生成的精子和卵子各隻含23對染色體中的一半,變成單倍性細胞,即配子。單倍體細胞精子和卵子的結合形成合子或稱受精卵,從而完成受精卵的形成過程。
B、 受精卵卵裂時期:此期是胚胎幹細胞(基因組)指數增長期。受精卵從輸卵管向宮腔移動的過程中,經曆了一係列快速的有絲分裂過程,此稱為卵裂。這一發育期開始於第一次有絲分裂,終止於胚泡的形成。受精卵的分裂基本上是一種典型的有絲分裂,其染色體的形態與結構和體細胞的相似,卵裂時期受精卵從2卵球到4卵球,再到8卵球,再到16卵球,再到32卵球……,再到桑椹期,最後到早期胚泡的形成。由此我們不難發現此期胚胎幹細胞(基因組)數量呈指數增長,出現加速增長的趨勢。受精卵卵裂過程的出現,一方麵表明其人體生物基因組的複製和表達過程呈現加速增長勢頭,另一方麵也表明其人體DNA含量呈快速增長勢頭,即呈幾何級數增長勢頭。
圖18 受精卵卵裂示意圖
C、 三胚層形成時期:此期是胚胎幹細胞(基因組)減速增長期。這個時期是胚胎細胞進行結構與功能的分化時期。胚胎幹細胞結構與功能分化時期也可稱為胚胎幹細胞的發育時期。其主要表現為胚胎幹細胞的分化與形態發生。此期胚胎幹細胞(基因組)的增殖開始轉化為減速增長,即負指數增長或稱對數增長,並且在結構與功能上出現多樣化的分化過程。同時有秩序地進行形態發生,首先形成三胚層。即經過這一階段的發育,形成了內、中、外三個胚層。外胚層形成了神經組織和表皮,中胚層形成了肌肉和結締組織,脈管係統和其他內髒器官,內胚層形成了消化管即上皮及其衍生化結構。三胚層的形成和分化決定了胚胎的中軸結構,形成了各個器官的原基,並為人體器官發生和組織分化打下了基礎。人體胚胎發育過程中三胚層的形成時期就相當於動物進化的原腸時期。盡管各種動物進化的原腸胚時期的變化與人體胚胎時期三胚層形成的變化差別很大,但它們的基本過程和發生機製是相似的。此期胚胎幹細胞數量的減速增長一方麵表明其生物基因組複製與表達過程開始趨緩,另一方麵也表明其DNA含量的增長出現拐點呈負指數增長或對數增長。
D、 胎兒形成時期:此期是胚胎幹細胞(基因組)增長的飽和時期。胎兒形成時期是人體完成胚胎發育過程的最後一個時期,是人體生殖分化的最後時期。我們知道,人體胎兒的形成需經過一定時間的發育,即三胚層結構與功能的分化。人體胎兒的形成過程將重演其進化的全部過程。胎兒的形成標誌著人體規範的確立。不同的生命個體在完成胚胎發育形成“胎兒”的曆程中,所達到的程度是不一樣的,例如:魚的胚胎發育隻能出現魚的幼體,蠑螈的胚胎發育隻能出現蠑螈的幼體,雞的胚胎發育隻能出現雞的幼體,豬的胚胎發育隻能出現豬的幼體,同樣,人的胚胎發育也隻能出現人的幼體(胎兒)。生物是進化發展的,不同的生命個體之所以在其胚胎發育過程中最終形成相應的幼體,主要原因是其受精卵內遺傳基因(縱向生命遺傳信息和橫向生命遺傳信息)所決定。這些基因中就包含著形成相應幼體的生命遺傳信息。生物體的生殖與發育過程就是這種遺傳信息的複製與表達過程。因此,不同進化層次,不同進化階段的生物,其胚胎發育所表達的性狀也是不一樣的。但高層次生物在表達的過程中,往往重演低層次生物的一些性狀,由此表明其在建立規範的曆程中,出現重演現象。參見德國學者E.H.海克爾生物發生律(Law of biogenesis)或生物重演律(law of recapitulation)。(例如:人體的胚胎發育過程就是整個生物係統進化的短暫而迅速的重演,這是一個以縱向遺傳信息表達為主的時期)
圖19 生物重演現象示意圖
此期胚胎細胞(基因組)的增長趨向於零增長狀態。最後出現增長的飽和現象。胚細胞的增長出現飽和現象,一方麵表明其生物基因組的複製和表達處於相對飽和狀態。另一方麵也表明其總體DNA含量的增長處於相對飽和狀態。

圖20 胎兒形成過程示意圖
4.1.2 生長發育階段
E、新生兒誕生時期:此期稱為人體各係統組織幹細胞(基因組)增長的延滯期。胎兒形成之後,通過分娩產生新生兒。新生兒的誕生標誌著人體作為一個規範的生命個體的正式形成並來到人間。這個時期,胚胎幹細胞發育過程可以看成已轉化為組織幹細胞發育。胚胎幹細胞的功能與結構的分化已經結束並形成新的生命規範體係。但此時是幼稚的,是不成熟的。各個組織係統的組織幹細胞還將要進行一次新的增殖。新生兒的誕生是一個非常重要的時期,這個時期可以看成是新與舊的交替時期,新生兒的誕生標誌著一個新個體生命時代的開始。
F、 青春發育期(組織幹細胞增殖時期):此期是人體各係統組織幹細胞(基因組)指數增長期。此期也可稱為人體生長的青春發育時期。新生兒誕生之後變成嬰兒,嬰兒初生時,牙齒、生殖器官等沒有長成,身體各種比例也和成人有差異,這個時期人體各係統內的組織幹細胞開始進行快速增殖並導致人體從幼體向成體發展。因此,此期也可稱為幼體生長期。幼體生長時期也是各係統組織幹細胞(基因組)增長的加速時期,出現各係統組織幹細胞(基因組)指數增長過程。這個時期的特點是不再形成新的器官,幼體器官隻是長壯,功能不斷加強並快速向成體發展。我們知道,人體生長是胚胎發育的繼續,新生兒時期的組織幹細胞具有很強的分生潛力,因此,幼體生長初期,各類組織幹細胞都能進行快速增殖。這就是所謂的胚後期發育。胚後期發育是指從母體生出幼體,它們與成體相比,無論在形態構造上,生理功能上以及生活習性上都存在一定的差別,新生兒誕生後還要繼續進行生長發育,人體出生後的生長發育過程從總體上講,同樣按加速方式發展,也就是人體總體的生長過程呈加速增長勢頭。但人體各係統、各器官的生長則以不同的速率,按不同的起止時間進行增長,這就導致人體生長過程出現異律現象。而異律現象的出現則表明各係統組織幹細胞生物基因組的複製與表達具有不同的增長速率。此期各係統組織幹細胞(基因組)從總體上看呈指數增長,由此充分表明其生物基因組的複製與表達處於加速增長狀態,同時也表明其DNA的總體含量的增長同樣處於加速增長狀態。
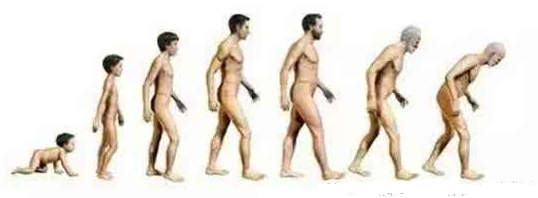
圖21 人體從新生兒到成年再到老年成長過程
G、 性成熟時期:此期是人體各係統組織幹細胞(基因組)減速增長期,也可以稱為組織幹細胞結構與功能特化時期。這個時期,各係統內組織幹細胞的增長開始趨向減速增長即負指數增長或稱對數增長。人體經過幼體長大為成體之後,經過發育過程,生殖係統達到功能成熟,這時人體的整個機體也完全趨於成熟,因此,稱這種發育過程為性成熟過程。人體性成熟的最主要內容是生殖係統完成發育,開始有了生殖機能。人體性成熟發育過程主要由內分泌係統和與生殖有關的部分出現了功能性變化。人體除了有雌、雄性別之外,同時個體還出現次生征或稱副性征。此期人體各係統組織幹細胞的增殖出現負指數增長,而這一切正表明其各係統內組織幹細胞生物基因組的複製與表達處於相對緩慢增長狀態,同時也表明其DNA含量的增長處於負指數增長狀態。
H、 衰老死亡時期:此期是人體各係統組織幹細胞(基因組)增長的飽和期。人體達到性成熟之後,在結構與功能上呈現種種衰退性改變,這種變化隨年齡而增加。最終導致人體的死亡。成熟機體的結構隨著年齡的增加而出現老化,稱為衰老。人體到了一定年齡就會停止生長,生長停止之後,經過一定時間就開始衰老。在這個時期,人體各係統組織幹細胞(基因組)的增長從總體上講趨向於零,即組織幹細胞(基因組)增長趨向飽和狀態,出現增長的飽和現象。衰老時期,身體構造和生理機能都在發生一係列的變化,如人到了中老年時期,毛發變白,皮膚變皺,牙齡脫落等等征狀。不過人體的每一個細胞有新生,也有衰老;有生長也有死亡,但壽命長短是不一樣的,隨著組織幹細胞的種類和環境條件的不同而出現差異。人體內組織幹細胞的衰老和死亡,雖然與整個機體的壽命密切相關,但二者並不完全一致。衰老的結果就是死亡,當機體的器官變得效率很低時,整個身體變得不能抵抗外界環境的變化,感染不易控製,體內的化學變化過程也失去了自我調控能力,最後,某一器官不再能執行其它器官賴以生存的功能,結果導致人體的死亡。此期各係統組織幹細胞(基因組)的增長處於相對飽和狀態,一方麵表明其各係統細胞生物基因組的複製和表達處於相對飽和狀態,另一方麵也表明其DNA的總體含量增長處於相對飽和狀態。
5.2 人體生命周期過程的動力學分析
5.2.1 傳統人們對人體發育生命周期的動力學的觀點
5.2.1.1馬爾薩斯的人口增長指數模型
早在馬爾薩斯之前,數學家歐拉(Leonard Euler,1707-1783)就提出過“歐拉的幾何人口增長的動態數學模型”。1748年歐拉在其出版的《無窮分析引論》第6章“指數與對數”中,所舉6個例子中第4個為:假設人口數量Pn關於年份n滿足方程Pn+1=(1+x)Pn(其中,n為整數,增長率x為正實數),若初始值為P0,則Pn關於n的表達式可以改寫為:Pn=(1+x)nP0,這即是“歐拉人口幾何增長動態數學模型”。
托馬斯·羅伯特·馬爾薩斯牧師(Thomas Robert Malthus,1766-1834)。英國生物統計學家兼經濟學家,出生於一個富有的家庭,他的父親丹尼爾是哲學家、懷疑論者大衛·休謨和讓·雅各·盧梭的朋友。馬爾薩斯年幼時在家接受教育,直到1784年被劍橋大學耶穌學院錄取。他在那裏學習了許多課程,並且在辯論、拉丁文和希臘文課程中獲獎。他的主修科目是數學。1791年他獲得碩士學位,並且在兩年後當選為耶穌學院院士。1805年他成為英國第一位(或許是世界上第一位)經濟學教授,執教於東印度公司學院。他的學生親切地稱呼他為“人口”馬爾薩斯教授。

圖22 馬爾薩斯(ThomasRobert Malthus,1766-1834)
1798年,馬爾薩斯出版了他的專著《人口論》又叫《人口原理》。此書從兩個不變法則出發:“食物為人類生存所必需;兩性間的情欲是必然的。論證人口以幾何級數(1,4,8,16,32,......)增加,生活資料以算術級數(1,2,3,4,5,6,......)增加,人口增長必然超過生活資料增長,人口過剩和食物匱乏是必然。因此,必然發生強大的妨礙、阻止人口的增加,這種妨礙就是貧窮與罪惡”。
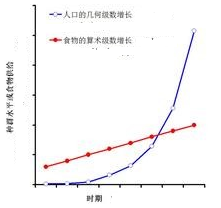
圖23 馬爾薩斯人口增長曲線圖
根據百餘年的人口統計資料,針對人口增長規律,提出了種群模型的基本假設:“在人口自然增長的過程中,淨相對增長率(即單位時間內種了、群的淨增長數與其總數之比)為常數r,以此為基礎,他從對人口增長和食物供求增長的分析中推導出了下述微分方程”。馬爾薩斯生物總數增長定律指出:“在孤立的生物群體中,生物總數N(t)的變化率與生物總數成正比”。其數學模型為:

其中r為常數。方程的解為:
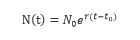
式中:N(t)為種群增長數量,N0為起初種群數量,t為時間,r為常數,e為自然數。
因此,遵循馬爾薩斯生物總數增長定律得任何生物都是隨時間按指數方式增長。在此意義下的馬爾薩斯方程,又稱指數增長模型。人作為特殊的生物總群,人口的增長也應滿足馬爾薩斯生物總數增長定律,此時的公式稱為馬爾薩斯人口方程。英國人口學家馬爾薩斯根據百餘年的人口統計資料,於1798年提出了人口指數增長模型。
5.2.1.2威爾霍斯特—珀爾方程
馬爾薩斯於1798年發表的《人口論》一書中提出人口增長的“指數規律”造成了廣泛的影響。1833年,威爾霍斯特以其著名的邏輯斯諦曲線描述人口增長速度與人口密度的關係,對其指數增長規律進行了修正。他把數學分析方法引入生態學。後來人們又把其邏輯斯諦(Logistic)曲線方程,稱之為威爾霍斯特方程。

圖24 威爾霍斯特(Pierre Francois Verhulst,1804-1849)
起初這個方程並未引起學術界的重視,到20世紀20年代被美國生物學家和人口學家珀爾等人重新發現,並用於分析人口增長過程。後來人們又將其廣泛運用於生物學、社會經濟增長的分析,特別是人體發育過程分析。因此,S曲線又稱珀爾生長曲線,又稱威爾霍斯特—珀爾方程。
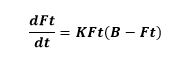
說明:用常微分dFt/dt=KFt(B-Ft)表示人體增長速度取決於對飽和極限的接近程度。B:人體增長的飽和極限;no:人體增長曲線的拐點年代;Ft:人體增長指標;t:時間;K:任意常數。
R.珀爾的生長曲線又稱珀爾曲線,是1938年比利時數學家威爾霍斯特(P.F verhulst)首先提出的一種特殊曲線。後來,現代生物學家珀爾(R.Pearl)和雷德(L·J·Reed)兩人把此曲線應用於研究人口增長規律研究。所以這種特殊的曲線又稱之為珀爾增長曲線,簡稱珀爾曲線。後來安全感人們又將其曲線應用於解釋人體和其它生物的生長發育過程的研究。

圖25 珀爾人體的S生長曲線
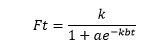
說明:Ft為人體增長量;t為時間變量(單位為年);k,a,b為大於零的常數;k為當t趨向於無窮大時的人體增長的最大值。
S曲線可分為個體生長曲線和群體(平均)生長曲線。一般是在橫軸上標出時間,縱軸上標出測定值。群體生長多呈S形曲線,這是最普通的生長曲線。從微生物直到人類的生物種群,其個體數的增加(人口增加),也常常符合此曲線。此曲線可分為兩種形態,即促進生長的前期和生長減衰的後期。兩種形態的轉折點(曲折點),動物相當於成熟期(青春期)。隨著動植物的種類和生長的時期以及器官的種類的不同,還可以得出另外的各種生長曲線,並能求出適合於這些曲線的方程式。生長過程也常可分為幾個生長曲線(指數曲線和S形曲線)。
斯柯孟(Scammon)根據珀爾曲線曾把人類生長的一般型(general type)描述呈S形曲線的生長型,見於身高、體重、體表麵積及其他外表的身體量度,與骨骼、肌肉、血液量、腎髒、消化、呼吸器官及其關聯部分。他將人體身高與體重為縱坐標,以人類生長年齡(時間)為橫坐標,得到的曲線叫生長曲線。後來人們經過長期研究發現這是一條被公認的生長曲線。
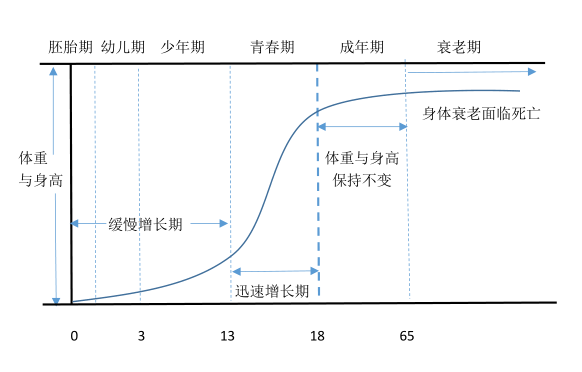
圖26 人體發育不同時期生長曲線
S生長曲線預測法也稱生長曲線模型(Growth curve models)是預測事件的一組觀測數據隨時間的變化符合生長曲線的規律,以S生長曲線模型進行預測的方法。一般來說,事物總是經過發生、發展、成熟三個階段,而每一個階段的發展速度各不相同。通常在發生階段,變化速度較為緩慢;在發展階段,變化速度加快;在成熟階段,變化速度又趨緩慢,按上述三個階段發展規律得到的變化曲線稱為生長曲線。綜上可見,R.珀爾曲線運用是十分廣泛,在生物學領域和社會經濟領域都有應用。但R.珀爾曲線是人們在考察人體個體發育和其它生物個體發育的上發現的生長曲線,它存在一定的局限性。可見曆史傳統上人們在考察人體的生長過程都是從身高和體重的變化上進行分析,因此得出人體生長過程遵循S曲線規律是必然的。這也是過去人們得出錯誤結論根源。
而我們則是從細胞和基因組擴增上考察,所以得出人體生長發育的雙S曲線理論。因此,從不同層次和不同角度看問題,得出的結論是不一樣的。所以,在解釋人體生命周期或其它靈長類生物等時可能要進行修正。因為人體或其它靈長類生物等有二次發育的過程。一次是在胚胎孕育時期,另一次是在出生後的生長發育時期。這兩個時期的增長是不一樣的,一個是胚細胞(胚胎幹細胞)的增長過程,另一個是體細胞(組織幹細胞)的增長過程。下麵我們就來分析它們是如何發育成長(增長)的。
5.2.2 現在的人體生命周期過程中細胞增長的動力學分析
現代細胞分子生物學告訴我們,人體細胞的增殖過程,可分為兩個階段,一是胚細胞增殖階段,二是體細胞增殖階段。這兩個細胞無論生長到那個階段。細胞核核酸含量是不變的,也就是細胞裏的DNA含量是不變的,這可以理解為其基因組是不變的。根據以上觀察,我們不難看出,人體的生命周期過程就是人體的生殖分化與生長發育過程。通過對人體胚胎幹細胞和組織幹細胞增長過程的細胞動力學進行分析,我們發現:
首先,人體從其受精卵(基因組)開始,一步步地進行分裂,即:一分為二,二分為四,四分為八,八分為十六,十六分為三十二,三十二分為六十四,……,最後到桑葚期;這個過程我們將其稱為人體胚胎幹細胞(基因組)的“加速增長期”或稱人體胚胎幹細胞的“指數增長時期”。之後到了三胚層時期,接著就是各種胚胎幹細胞進行結構與功能分化,有的胚胎幹細胞開始分化形成大腦神經細胞、有的胚胎幹細胞分化形成四肢骨骼細胞、有的胚胎幹細胞分化形成內髒細胞、有的胚胎幹細胞分化形成各種血管組織細胞……等等。隨著胚胎幹細胞分化過程的進行,這時的胚胎幹細胞(基因組)數量的增長開始趨向緩慢,出現“負指數增長”即“對數增長”,最後形成“胎兒”。出現人體胚胎幹細胞(基因組)增長的“飽和現象”,並因此構成人體胚胎幹細胞(基因組)增長的邏輯增長趨勢。由此我們可以清楚地看到人體完成了第一個“邏輯增長過程”。出現第一個“S型增長曲線”。人體在進行生殖與發育過程時,第一階段進行的是胚胎幹細胞(基因組)的增殖,從細胞裂變時起,進行指數增長時總是會出現拐點的。假如不出現拐點,那麽,胚胎幹細胞(基因組)就會不斷地指數增長下去,其生物基因組的複製過程就會趨向無窮大,最終會出現什麽結果呢?它必然會出現巨大的胚胎幹細胞體(基因組複合體)。並且你可以想象它長多大就可以有多大。然而,這是一種荒唐的結果。實際上是不可能出現的。從正常的人體生殖與發育過程上看並沒有出現上述情況,而是胚胎幹細胞隨機性地進行了結構與功能的分化與調整。通過生物發生過程最終形成了人體“胎兒”。因此,出現拐點是必然的,是一種邏輯的選擇。正是因為這種選擇導致人體生長的第一個S曲線增長過程的出現。
然後,隨著“新生兒”的誕生,新的人體生物規範的確立,人體又向一個新的階段開始邁進。這時人體內各個係統在新規範的控製和調節之下,各個係統組織內的組織幹細胞(基因組)又分別開始進行增殖生長,起初其增長速度是緩慢的,但到了青少年“青春發育時期”,人體各大係統組織內的組織幹細胞(基因組)增長速度又開始明顯加快,出現加速增長勢頭,呈“指數增長”。這也是一個人個體發育過程中出現的“典盛時期”。這個時期的出現意味著人體作為一個規範的生命係統進入了一個加速增長時期。這個時期人體的神經係統、內分泌係統、骨骼係統、運動係統、呼吸係統、免疫係統等等均進入了一個最佳狀態。隨著這個時期的結束,人體又開始進入到一個發育成熟狀態,出現性成熟。各係統內的組織幹細胞(基因組)的增長開始進入“負指數增長”時期,即出現對數增長期。各係統內的組織幹細胞出現特化現象。隨後,隨著人體向中老年發展,各係統內的組織幹細胞(基因組)增長又開始趨向於“零增長”,進入飽和狀態,各係統內的組織幹細胞出現老化和退化現象。最後人體開始走向衰老消亡。由此我們不難看到,人體從“新生兒”誕生時起一直到衰老消亡,這個階段又完成了一次“邏輯增長過程”,出現第二個“S型增長曲線”。第二階段進行的是人體各係統組織內組織幹細胞的增殖,組織幹細胞的增殖起初也是一個指數增長的過程。然而,組織幹細胞的增殖也不可能長期指數增長下去,如果隻增殖不分化、不特化,體細胞的增長也將趨向無窮大,最後這個人體就會長成為一個巨大的生物體,並且是沒有進行有效分化、有效發育的生物體,同時也是一個青春常在,發育不全的生物體。你也可以想象他能長多大就可以長多大,這種結果也是不存在的,同樣是荒唐的。因為人體從幼體新生兒時起就開始生長發育並向成體轉化,進行了人體結構與功能的分化,最終導致性成熟,同樣也出現了組織幹細胞(基因組)增長的拐點,出現負指數增長並符合邏輯地向中老年方向發展,最後衰老消亡。從而導致人體生長第二個S曲線增長過程的出現。
我們把第一次完成的S型曲線增長過程稱為非常規增長過程,而將第二次完成的S型曲線增長稱為常規增長過程,前者沒有規範,後者有規範;前者沒有特異性,後者有特異性;前者胚胎幹細胞分化是隨機性的,後者組織幹細胞分化則是確定性的。這兩個增長時期的結合構成人體全部生命周期過程,而人體整個生命周期過程則表現為一個連續的“雙S型增長曲線”(如圖2所示)由此完成人體從“受精卵子”開始到“胎兒”形成再從“新生兒”誕生到最後“衰老消亡”整個生命周期的全部過程。

圖27 人體生長過程中細胞(基因組)增長的雙S曲線規律示意圖
說明:人體的生長過程包括兩個階段:
第一階段:人體胚胎幹細胞(基因組)增長階段(生殖分化階段:A期、B期、C期、D期);表現為第一個S型曲線:(以縱向遺傳信息表達為主的增長動力過程)
第二階段:人體組織幹細胞(基因組)增長階段(生長發育階段:E期、F期、G期、H期);表現為第二個S型曲線。(以橫向遺傳信息表達為主的增長動力過程)
所以,人體生長過程中,其總體細胞(基因組)增長遵循連續的雙S曲線規律。我們分別對其兩次S曲線增長過程(胚胎幹細胞增長與組織幹細胞增長過程)進行積分,然後再對兩次積分相加求和,最後得到人體生長過程中全部細胞(基因組)增長的總量。其積分和數學表達式(動力學模型)如下:

式中:Ft人體細胞(基因組)增長總指標;Fta胚胎幹細胞(基因組)增長指標;Ftb組織幹細胞(基因組)增長指標;t人體細胞(基因組)增殖總時間;ta=t0----t1胚胎幹細胞(基因組)增長時間;tb=t1----t2組織幹細胞(基因組)增長時間;K1胚胎幹細胞(基因組)增長係數;K2組織幹細胞(基因組)增長係數;M胚胎幹細胞(基因組)增長的飽和極限;N組織幹細胞(基因組)增長的飽和極限;n1胚胎幹細胞(基因組)增長曲線的拐點;n2組織幹細胞(基因組)增長曲線的拐點;
5.2.3 生物增長理論的突破與發展
總之,我們不難看出“生命周期雙S曲線規律”是對“歐拉-馬爾薩斯指數增長模式”和“威爾霍斯特-珀爾S曲線增長模式”的突破與發展。它的發現具有重要的生命科學意義和價值。正像“歐拉-馬爾薩斯指數增長模式”和“威爾霍斯特-珀爾S曲線增長模式”可以廣泛地應用到經濟和社會發展的研究一樣,“生命周期雙S曲線規律”也同樣可以應用到經濟和社會發展的研究之上。而這一切將改變過去人們的看法。
6人類基因組生命遺傳信息的複製與表達對人體生殖與發育過程的深刻影響
通過對人體生殖與發育過程中細胞(基因組)增長的係統分析與研究,我們發現人體生殖與發育過程遵循生命周期雙S曲線規律。然而,人體的生殖與發育過程遵循生命周期雙S曲線規律的分子生物學依據是什麽?人體生殖與發育過程中其基因組遺傳信息的複製與表達和人體生殖與發育過程的關係是什麽?為什麽人體的生殖與發育過程一方麵表現出非特異性和非規範性,另一方麵又表現出特異性和規範性?對此我們將從以下幾方麵來加以闡述:
6.1 人體生命周期雙S曲線規律的細胞與分子生物學基礎
我們認為,人體的生命周期過程就是人體生物基因組遺傳信息的複製與表達過程,並且這個過程遵循雙S曲線規律。在我們看來,人體生物基因組遺傳信息的複製與表達,主要是分二個階段來完成:
首先,形成幼體胎兒。受精卵(基因組)進行裂變增殖(指數增長),也就是胚胎幹細胞進行基因組的大量快速複製,與此同時進行一些非特異性遺傳信息的表達,即縱向生命遺傳信息(一般共同信息,即種係發育信息)的表達,完成生物發生過程(重演生物進化曆程)。因為縱向遺傳信息是大家共有的,所以不具有特異性,人人基本相同。這種非特異性的遺傳信息的複製與表達,使得人體胚細胞在增殖與發育的過程中出現結構與功能的分化,經過生物發生過程,即從受精卵到三胚層,最後導致胎兒的大腦、四肢和內髒等器官的出現,形成幼體胎兒,完成第一次胚胎幹細胞的增殖與發育或人體生物基因組的複製與表達過程。此次胚胎幹細胞的增殖過程是沒有特定方向的,是一種隨機性的增殖與分化過程,胚胎幹細胞在進行細胞的分化過程中,具有全能性,即任何一個胚胎幹細胞都可以分化成人體胎兒的大腦、四肢和內髒器官。因此也可以說是一種非確定性的發育分化過程。此階段人體生物基因組的複製與表達過程遵循第一個S型曲線。
其次,形成成熟個體。新生兒誕生後,人體各係統內的組織幹細胞進行裂變增殖(指數增長),也就是各係統組織幹細胞進行生物基因組的大量快速複製,與此同時則進行特異性遺傳基因信息的表達,即一些橫向生命遺傳信息(特殊差異信息,即個體遺傳信息)的表達。完成人體特異性的生長發育過程(重現親代的一些遺傳特征)。橫向遺傳信息不是大家共有的,是具有一定的種族、家族、個體等差異性和專有性,因此具有一定的特異性。這種特異的遺傳信息的複製與表達使人體各係統的體細胞在增殖與發育的過程中進行了結構與功能的分化或特化,最後導致人體成體的形成,使人體從幼體轉化為成體,隨後衰老消亡。完成第二次人體基因組的複製與表達過程。此階段,組織幹細胞的增殖與分化過程是有特定方向的,是一種確定性的、非隨機性的增殖與發育過程。因為人體組織幹細胞的增殖與發育過程是沿著特定的方向進行的,不同係統組織的組織幹細胞的分化是按照各自係統組織細胞的發育方向進行的。表現出發育過程的專業性。此階段人體遺傳基因組的複製與表達過程遵循第二個S曲線。
人體生物基因組的複製與表達在這兩個階段均出現了加速增長期和增長的飽和期,但這兩個階段細胞的增殖與發育所表現出的特征是不一樣的。從上述分析中我們可以看出,第一個時期是胚胎幹細胞非規範性增殖與發育(非特異性增殖與發育);第二個時期則表現為組織幹細胞規範性增殖與發育(特異性增殖與發育)。這兩種細胞的增殖與發育過程構成了人體整個生命周期過程,並遵循雙S曲線規律。由此我們不難看出,人體生物基因組的複製和表達是周期節律性的,並且是按照一定的程序進行的。即人體從受精卵開始,通過受精卵的裂變過程,使其胚胎幹細胞生物基因組進行了大規模複製,導致人體生物基因組數量的大規模增長,也就是胚胎幹細胞的大量增殖。這種胚胎幹細胞大量增殖的意義是十分重要的,其為以後胚胎幹細胞的隨機分化做好了充分準備。從海克爾重演律上講,人體的生物發生過程(生殖與發育過程)就是生物係統進化過程短暫而迅速的重演,人體在生長發育過程中,其生物遺傳基因組的複製與表達並不是一個漫長的演化過程,而是在其胚胎時期,就將整個生物係統的進化過程進行了一次重演。生物進化最本質的方麵是生物基因組的進化。生物從低級到高級,從簡單到複雜,從低層次到高層次,主要區別在於其生物基因組結構序列的變化。因為,人體的生命遺傳信息全部貯存於人體生物基因組上。我們知道,人類生物基因組圖譜的形成,從縱向上看是從生物界一步步地進化而來的,但從橫向上看,則又是親代父係與母係染色體雜交後形成受精卵而獲得。也就是一方麵,人體生物遺傳基因既然是從生物界進化中獲得,就必然包含有生物進化過程的生命遺傳信息,因此,在其胚胎發育過程中就要逐步進行複製與表達。同時,從另一方麵看,人體的遺傳基因又來自於親代的染色體雜交,因此也同樣包含有親代的生命遺傳信息,這些生命遺傳信息也要複製與表達,這點在人類個體胚後發育時期得到很好的體現。

圖28 2000年沈律先生參加“第五屆海內外生命科學論壇”並在會上宣讀論文《生物進化與生物發生的基本規律——對人類基因組的形成及其複製與表達過程的係統研究》(參見《幹細胞與發育生物學》(北京:軍事醫學科學出版社,2000年.217-229)。客觀的說,這篇論文的宣讀與發表標誌著人們對生命進化規律的認識進入了分子水平(基因組)新階段(它不同於R.珀爾的個體發育和群體增長階段),這正好與華生和克裏克的工作(發現DNA和RNA雙螺旋結構)銜接為一體。它是從分子生物學水平用生物計量學(生物數學)方法闡述了生命的實質及其進化發育規律。第一,從孟德爾遺傳定律到摩爾根遺傳定律再到遺傳因子(基因)矩陣分布定律,這些規律都是一脈相承的,它們都是用排列和組合的方式一個比一個更全麵深入地了解生物遺傳基因在個體和群體的分布情況,從而揭示了生命遺傳、發育和進化的本質聯係。第二,從歐拉-馬爾薩斯指數增長理論到威爾霍斯特-珀爾增長理論再到雙S曲線規律理論,它們也是一脈相承的,而生命周期雙S曲線規律則是對歐拉-馬爾薩斯指數增長理論與威爾霍斯特-珀爾增長理論的突破與發展。因此,它們在生命科學的意義上與前人的工作是同等重要的。
6.2 人體生命周期過程中其細胞的增殖與分化和其基因的複製與表達之間的相互關係
我們認為在人體生命周期過程中,人體細胞的增殖與分化和人體生物基因組的複製與表達直接相關。現代生物學發現,人體生物基因組序列在其整個生命周期中或整個細胞增殖與分化過程中,它的結構序列是不變的。也就是人類個體發育從受精卵到胚細胞再到體細胞其基因組序列是恒定的。而不同時期其分化發育出來的生物性狀則又不一樣,這一切主要取決於人體生物基因組二次大規模複製與表達。我們發現,人體細胞二次大規模增殖與分化過程和人體生物基因組二次大規模複製與表達過程直接相關。通過對人體生殖與發育過程的係統分析,我們發現人類個體在其發育過程中出現了兩次大規模生物基因組的快速複製與表達,前者在受精卵(胚胎幹細胞)裂變時期,後者在各係統組織幹細胞裂變時期。這兩個時期其細胞數量均出現了指數增長勢頭,這兩次快速細胞增殖的意義何在呢?
第一次裂變增殖導致細胞快速增長,同時分化發育成胎兒,這個時期其胚胎幹細胞數量的快速增長意味著人體生物基因組的複製出現了幾何增長(指數增長)勢頭,也就是人體生物基因組的數目呈快速增加。隨後,這些大量快速複製的人體生物基因組又迅速地將某些基因片段進行表達,使胚胎幹細胞隨機分化成了胎兒的大腦、四肢、內髒等器官,在這一過程中,表達的由於是縱向的生命遺傳信息(一般共同信息,即種係遺傳信息),沒有明顯的個體差異;差異性,特異性不明顯。沒有什麽排他性,排斥性。是一種非規範信息表達過程。人體胚胎發育過程可以看成生物長期進化的短暫迅速重演的過程。參見德國學者E.H.海克爾生物發生律(Law of biogenesis)或生物重演律(law of recapitulation)。
第二次細胞裂變增殖也同樣導致各係統組織幹細胞的快速增長,其生物基因組數目第二次出現了幾何增長(指數增長)勢頭,與此同時各係統生物基因組中的某些基因片段也進行了功能表達,而在這一過程中,主要表達的是橫向的生命遺傳信息(特殊差異信息,即個體遺傳信息),因此出現了很大的個體差異。橫向信息表達具有強烈的特異性和排他性。是一種特征性表達,是一種特別性表達,是一種規範性信息表達過程。
兩次人體生物基因組的複製與表達正好構成了人類個體生殖與發育的全過程。由此我們發現人體在其細胞的增長的過程中,盡管其生物基因組結構序列不變,但其生物基因組數量則增加了;細胞的分化是其生物基因組中的某些基因片段的功能表達,但不同時期所表達的遺傳信息又不一樣。人體生物基因組的複製與表達是相繼進行的。複製導致細胞數量的增長,而表達則導致細胞結構與功能的分化。人類的個體發育過程一方麵可以看成是細胞的增殖和分化過程的結合,另一方麵也可以看成是其生物基因組複製與表達過程的結合。在人體的整個生命周期過程中,其細胞數量的增長與細胞結構的分化並存,其生物基因組的複製與其生物基因片段中生命遺傳信息的表達並存。同時其細胞數量的增長與其生物基因組的複製相關一致,其細胞的功能分化與其生物基因組遺傳信息的表達相關一致,沒有人體生物基因組的複製就沒有人體細胞數量的增長,沒有人體生物基因組遺傳信息的表達就沒有人體細胞的結構與功能分化。這兩者是密不可分的。人體生物發生的生命周期雙S曲線過程正是這兩個方麵的真實反映。
6.3 人體生命周期過程中不同時期產生非特異性、非規範性和特異性、規範性的機製
我們認為,人體生殖與發育過程既表現為一定的非特異性、非規範性,又表現為一定的特異性、規範性。我們發現,人體細胞在從胚細胞增殖發育向體細胞增殖發育轉化的過程中,其特異性和規範性也在不斷加強,並且表現出質的變化。例如,人與人之間在其胚胎時期,他們的免疫排斥反應是不明顯的,但一旦到了體細胞增長時期,情況也就不一樣了,其表現出強烈的免疫排斥反應。其原因何在呢?現代醫學免疫學發現這主要是因為人體體細胞膜上出現了一種組織相關性抗原,它是一種蛋白質,而這種蛋白質的合成就是靠一種特殊遺傳基因調控的。這種組織相關性抗原導致人體特異性的產生。但同卵兄弟或姐妹則不出現上述排斥反應,其原因何在呢?這主要是因為同卵兄弟或姐妹的受精卵基因組是完全一樣的,並且在複製與表達的方式上,時間節律上也是一樣的,同時又是以相同的方式進行胚胎幹細胞和組織幹細胞增殖發育。異卵兄弟或異族兄弟則具有上述排斥反應。這表明,在同一個家族,不同時期受精的受精卵其生物基因組的結構序列是不一樣的,因此其遺傳基因的複製與表達方式也就不一樣。不同家族的人群其受精卵的生物基因組結構序列更是不一樣,因此其遺傳基因功能表達更具有特異性和規範性。通常情況他們在排斥反應的強度上表現為:同卵兄弟小於異卵兄弟,異卵兄弟小於異族兄弟。這一切產生的根源就在於人體生物基因組在形成的過程中,由於生物遺傳基因的自由分離與自由組合而導致人體不同生物基因組結構序列的形成。所以我們不難看出,不同的排列與組合產生不同的生物基因組圖譜,不同的生物基因組圖譜導致不同的生物性狀的產生即不同的規範或不同的特異性產生。因此,生物基因組圖譜的差異性及其功能表達的差異性是產生特異性排斥反應的關鍵。同種胚胎幹細胞之所以表現為非特異性,主要原因是他們的生物基因的非特異性表達,所謂非特異性表達是指胚胎幹細胞遺傳信息的表達過程隻是啟動了一些共同的生物基因片段或一般基因片段,未啟動不同的基因片段或特殊基因片段,合成的蛋白質沒有特異性。因此,胚胎幹細胞發育分化過程表現為非特異性和非規範性。這點從遺傳基因信息的表達上看,主要是由於在第一個S型曲線增長期,人體遺傳基因主要表達的是縱向的生命遺傳信息(一般共同信息,即種係遺傳信息),而縱向生命信息是每個人都共有的。因此不具有特異性。但到了組織幹細胞時期,組織幹細胞基因表達的主要是一些橫向的生命遺傳信息(特殊差異信息,即個體遺傳信息),這些遺傳信息不是人人都一樣的,具有種族、家族和個體的差異性和特殊性,因此,組織幹細胞的發育與分化過程表現出特異性和規範性。也就是說,到了組織幹細胞增長時期,隨著特殊基因片段的不斷啟動、不斷表達,並且將特殊基因信息都表達了,合成的蛋白質自然具有很強的特異性。這樣,個體免疫排斥反應也就更加強烈了,個體特異性、差異性也就更加明顯了。還有,在人體的生命周期過程中,胚胎幹細胞進行結構與功能的分化完全是一種隨機的過程,是一種隨機的發育分化過程,沒有規範的約束。那些細胞分化發育成神經細胞,那些細胞分化發育成內髒細胞,這些都是隨機的,不確定性的。但到了組織幹細胞結構與功能的分化發育時期,情況就不一樣了,組織幹細胞的發育分化過程是嚴格按照其既定的細胞類型進行的。神經細胞隻能向神經細胞進行發育分化,肝細胞隻能向肝細胞進行發育分化。不是隨機性的,而是一種確定性的發育分化過程,是有規範約束的。這一點對我們理解人體生殖與發育過程的非規範性和規範性也是非常重要的。
總而言之,我們認為,人體生命周期過程從本質上講是人體生物基因組遺傳基因的複製與表達過程。在這一過程中,人體生物遺傳基因組的複製與表達是有周期節律性的。在我們看來,人體生物遺傳基因組的複製與表達具有兩個指數增長期和兩個增長飽和期。它們分別發生在人體受精孕育階段和人體生長發育階段。而這兩個階段的出現與人體胚胎幹細胞增殖分化和組織幹細胞的增殖分化是直接相關的。同時也與人體生物遺傳基因組的複製和非特異性(非規範性)、特異性(規範性)的功能表達直接相關。前者表達的是縱向生命遺傳信息(一般共同信息,即種係遺傳信息),後者表達的是橫向生命遺傳信息(特殊差異信息,即個體遺傳信息)。兩者的結合構成了人體生殖與發育的全過程。並且這個過程從總體上講遵循生命周期雙S曲線規律。
6.4 人類基因組遺傳基因表達的睡眠理論
現代生命科學研究表明,基因表達是指細胞在生命過程中,把儲存在DNA序列中遺傳信息經過轉錄和翻譯,轉變成具有生物活性的蛋白質分子。生物體內的各種功能蛋白質和酶都是由相應的結構基因編碼信息表達完成的。人類基因組遺傳基因表達一是形成奢侈基因,二是形成管家基因。這兩類基因的表達約占整個基因組的5-10%。分子雜交等大量實驗表明,在人體細胞的全套基因組中,隻有少數基因(5-10%)表達。基因組中表達的基因分為兩類:一類是維持細胞基本生命活動所必須的,稱管家基因,如各種組蛋白基因。另一類是指導合成組織特異性蛋白的基因,對分化有重要影響,稱奢侈基因,即組織特異性表達的基因,如表皮的角蛋白基因、肌肉細胞的肌動蛋白基因和肌球蛋白基因、紅細胞的血紅蛋白基因等。這類基因與各類細胞的特殊性有直接的關係, 是在各種組織中進行不同的選擇性表達的基因。管家基因是維持細胞生存不可缺少的,奢侈基因和細胞分化有關,是組織特異性表達有關的基因,在特定組織中處於非甲基化或低甲基化狀態,而在其他組織中呈甲基化狀態。幾乎所有的甲基化過程均發生在二核苷序列5'-CG-3'中的C上。使胞嘧啶變為5'-甲基胞嘧啶。而含有這種甲基化CG的序列,對應於染色體上的兼性異染色質區域。管家基因以組成型方式在所有細胞中表達,而奢侈基因在特定組細胞中得到表達。這些基因的特異表達與否,決定了生命曆程中細胞的發育、分化、細胞周期的調控、體內平衡、細胞衰老、甚至於程序化凋亡。對不同類型,不同分化時期細胞的基因或基因表達情況的研究,可以獲得整個細胞生命過程的信息。細胞在不同自然或人工理化因子作用下代謝過程變化甚至於病變,基因也將選擇性表達。
我們認為,人體發育過程可分為非常規發育和常規發育兩個過程,前者是一般生物遺傳信息的表達,後者是特殊遺傳信息的表達。人體整個發育過程就是把這兩類基因信息全部表達的過程。人在胚胎時期主要表達的是縱向基因信息,而出生後主要表達的是橫向基因信息,這兩組信息全部表達後,這個人也就對束了全部生命過程。現在人們認為人類基因組基因信息隻表達了5-10%奢侈基因和管家基因。而我們認為人類基因,被人們稱之為垃圾基因90-95%是在胚胎發育時期就完成了表達。因此,可以說人體發育的整個過程,100%基因都得到表達。因為人體整個發育過程進行了一次生物進化過程的重演。基因隻是在不同時期是按照一定的程序在進行表達,是有秩序的在表達。隻不過是表達完之後,就處於睡眠狀態或休眠狀態。之後需要的時候再去表達。這種需要有正常需要和異常需要兩種。正常需要是在適當的時期,人體發育過程的生理需要,而異常需要則是病理狀態的需要,從而導致人類遺傳疾病的產生。由此,我們提出人類基因組表達的睡眠理論。這個理論是建立在生物基因組遺傳信息矩陣分布定律和人體生命周期雙S曲線規律基礎上的。沒有上述規律的認識,這個理論就失去了存在的基礎。人類基因組遺傳基因信息的表達與人體發育過程是非常密切的,人體發育過程有正常發育過程與異常發育過程。就看基因是不是在合適的時間進行表達。如果是,那就會導致正常發育;如果不是,那就會引發遺傳疾病。人類基因組基因表達過程,既有自我修複功能,又有自我關閉功能。它們能在一定的時間內實現自我關閉和自我修複。在不同的時間內,人類基因組遺傳基因信息表達。例如:如果在胚胎時期出現異常,就會引發發育個體出現畸變,產生畸形兒;如果在出生後出現異常表達,就會引發各種遺傳性疾病,甚至出現腫瘤。從本質上講,一個是胚胎幹細胞發育出現的問題,另一個是組織幹細胞發育出現的問題。人類基因組基因信息的表達往往是一有種間斷性的,不同時間基因表達是有規定的。有的基因處於表達狀態,而有些基因則處於睡眠狀態。
7 結 論
綜上所述,我們首先對生物新陳代謝一般均衡定律進行分析,了解生物代謝同化與異化過程的相互關係,提出新陳代謝指數概念;其次對生物基因組的形成及其生物遺傳信息中心法則進行分析和介紹,提出生物基因組的形成是通過縱向生物遺傳信息和橫向生物遺傳信息的雜交而成。人類基因組包含著兩類生物遺傳信息,一類是縱向生物遺傳信息,另一類是橫向生物遺傳信息,而這兩類生物遺傳傳信息則是通過生物遺傳信息中心法則進行傳遞。通過對生物遣傳定律進行進一步介紹,我們發現生物第四遺傳定律是一個非常重要的生命規律。通過對生物基因組遺傳信息矩陣分布特征的研究,發現不同的生物隨著其基因組縱向生命遺傳信息(一般共同信息,即種係遺傳信息)和橫向生命遺傳信息(特殊差異信息,即個體遺傳信息)質與量的不同,而表現出不同的矩陣分布狀態。並且從低級到高級,從簡單到複雜遵循一定的分布規律。各種生物隨著其生物進化和生物發生層次的不同而處於不同的時空發育過程中,並表現出一定的矩陣分布狀態。這種矩陣分布狀態是每個生物所特有的。據此,我們將其確定為“生物進化的遺傳信息矩陣分布定律”。根據這個定律,我們可以了解任何一種生物的進化程度和發育分化程度。第三,通過對人體生物發育過程的研究,我們發現,在人體生殖與發育過程中,人體進行了兩種細胞的增長。前者是胚胎幹細胞增長過程,後者是各係統組織幹細胞的增長過程。前者表現為非規範性和非特異性增長,後者則表現為規範性和特異性增長。這兩個過程結合表現為雙飽和曲線(雙S曲線)。因此我們認為人體的生殖與發育過程遵循“生命周期雙S曲線規律”。這就是我們通過對人體生殖與發育過程的係統觀察及其細胞增長的動力學分析所得出的結論。同時我們認為,根據海克爾重演規律,人體的生殖與發育過程是整個生物係統進化發展過程的一個短暫而迅速的重影。因此不僅人類個體的生物發育過程遵循生命周期雙S曲線規律,而且整個人類乃至整個生物係統的進化與發展過程也應遵循生命周期雙S曲線規律。最後,通過對這兩個規律進行相關性研究,我們發現,人體生殖與發育過程之所以遵循生命周期雙S曲線規律,其根本原因是在人體生物基因組中包含兩類生命遺傳信息,一是縱向生命遺傳信息(一般共同信息,即種係遺傳信息),另一是橫向生命遺傳信息(特殊差異信息,即個體遺傳信息);這兩類生命信息的複製與表達導致人體生殖與發育過程遵循生命周期雙S曲線規律。因此,我們認為,加強對生物新陳代謝一般均衡定律、生物進化的遺傳信息矩陣分布規律和生物發育的生命周期雙S曲線規律及其相關性研究,即加強對人類生物基因組的形成及其生命遺傳信息(縱向生命遺傳信息和橫向生命遺傳信息)的複製與表達過程的係統研究,這將有助於我們從根本上最終揭示生物遺傳、進化與發育的基本規律。
參考文獻
1984.黃厚哲編著.生物學概論.北京:高等教育出版社
1997.陳閱增主編.普通生物學.北京:高等教育出版社
1980.鄭國錩編著.細胞生物學.北京:人民教育出版社
2007.翟中和,王喜忠,丁明孝主編.細胞生物學.北京:高等教育出版社
2007.鄭集,陳鈞煇編著.普通生物化學.北京:高等教育出版社
2006.戴灼華,王亞馥,粟翼玟.遺傳學.北京:高等教育出版社
1999.徐克學著.生物數學.北京:科學出版社,
2015.趙斌著.生物數學簡史.北京:中國科學技術出版社
2008.王凱捷,李勇智著高等數學.北京:高等教育出版社
1985.袁蔭棠編.概率論與數理統計.北京:中國人民大學出版社
1972.C.達爾文著.物種起源.北京:科學出版社
1974.N·海克爾著.宇宙之謎.上海:上海人民出版社
1964.B·H·威利爾等主編.發育分析.北京:科學出版社
1982.K·L·穆爾著.人體發生學.北京:人民衛生出版社
1963. B·M·派登著.人體胚胎學.上海:上海人民出版社
1981.M.S.Thalr編著.醫學免疫學.北京:人民衛生出版社
1994.Scott F.Gilbert.Developmental Biology.Sinauer Associates,Inc.
1989. C.Starr.Biology.fifth edition.Belment:Wadsworth publishing company
1981.W.R.Atchley.Evolution and speciation.London:Cambridge university press
1985.M.W.Strickberger.Genetics.third edition.NewYork:Macmillan rublishing company
http://blog.sciencenet.cn/blog-38450-1067806.html
上一篇:現代數學概述
24 武夷山 徐令予 彭真明 史曉雷 秦逸人 趙克勤 楊波 楊正瓴 姬揚 王德華 王從彥 蔡小寧 李穎業 陳楷翰 檀成龍 郭新磊 尤明慶 劉煒 汪曉軍 趙斌 魏焱明 李勝文 wangbin6087 scientist
發表評論 評論 (13 個評論)
- [13]李勝文
- Your Chinese abstract sounds great, but its English should be improved for the clarity, the flow of logic, and the cohesive narrative.
- [12]李勝文
- Superb! I'd ask "what's the driving force of life?" Newton and Einstein explain how planets move/ things fall in place, but never before anyone shows how the driving force of life works. Its origin? Can AI drive itself? Can organic life compete with AI life? Another aspect of your theory was how pathological conditions kick in? fascinating! Thanks so much for your message as I didn't know your stuff until you tell me. Can you model cancer? It'll save a lot of lives!
- [11]魏焱明
-

excellent! 可以寫成專著了!
- [10]郭新磊
-
- [9]張學文
- 在建立半定量的學說?!
學習。
我猜目的性(某個體現目的量試圖極大化)應當是生物活動的指標。 - 博主回複(2017-7-26 06:29):謝謝您的評論,是的,生物的目的性就是一方麵保持自身係統的相對獨立與穩定,另一方麵保持與外界動態平衡。這兩個方麵就是建立一般均衡理論的核心內容。生物的新陳代謝、遺傳、進化、發育等等生命過程中無不體現著這種相對平衡。這種相對平衡一旦被打破,就會出現異常,甚至是疾病。但生命過程中一旦出現異常也會出現自我修複和自我修正的過程。不能修複,那就要靠醫學治療來解決了。
定量化(計量化)分析生命現象,是未來生命科學發展的趨勢,生命科學隻有真正走上計量分析的軌道,才能走向成熟。才能成為一門真正意義上的成熟科學。才能成為真正意義上的21世紀帶頭學科。
我的這篇論文隻是在生命科學計量化方麵做出的一點小小嚐試,今後還有很多大量的工作問題有待人們去解決。論文中肯定存在有這樣那和樣的問題,還請張老批評指正。
- [8]蔡小寧
-
- 博主回複(2017-7-26 04:41):謝謝!
- [7]楊正瓴
-
- 博主回複(2017-7-26 04:42):謝謝!
- [6]wangbin6087
- 其實計算也是分層次的。生物分子由小分子構成、小分子由原子構成、原子由基本粒子構成、基本粒子由量子構成(這個觀點大家現在還難以接受,因為量子是科學不能觀測的層次故在現象之外,科學隻能觀測量子的集合所呈現的現象),所以在不同的層次會有不同的算法和計算。真正的本底計算在真空量子的層次,量子糾纏就是真空的性質。
- [5]wangbin6087
- 博主回複的十分精彩。從中可以感悟到這樣的道理:我們在現象(形式)層次看到的演化,在本質層次來說就是計算。形式是本質的表達、演化是計算的表達。用4樓趙克勤老師的概念說,演化和計算就是形式與本質對立統一的集對關係。
- [4]趙克勤
- 入《讀博記語--生命在於計算》http://blog.sciencenet.cn/blog-329317-1067973.html
- 博主回複(2017-7-25 03:01):謝謝您的大力支持和宣傳。
- [3]張義國
- 蝌蚪士全文轉發你的工作,請把word版文稿寄給kedoushi@163.com
- 博主回複(2017-7-26 04:49):我已經看到我的論文。
- 博主回複(2017-7-25 03:00):謝謝您的大力支持和宣傳。在你們的《蝌蚪士》上,我怎麽能閱讀到我的這篇文章?
- [2]wangbin6087
- 如果是這樣,建議你更應該思考一下計算主義。DNA計算機這一概念的提出和實驗證明,會為你的努力提供一個清晰的方向和方法參考。
- 博主回複(2017-7-25 02:59):DNA計算機是一種生物形式的計算機。它是利用DNA(脫氧核糖核酸)建立的一種完整的信息技術形式,以編碼的DNA序列(通常意義上計算機內存)為運算對象,通過分子生物學的運算操作以解決複雜的數學難題。
機器計算的曆史可以追溯到1641年,當年18歲的法國數學家帕斯卡爾成功地製造了一台齒輪傳動的八位加法計算機。這使人類計算方式、計算技術進入了一個新的階段。後經過人們數百年的艱辛努力,終於在1945年成功地研製出了世界上第一台電子計算機。從此,人類進入了一個全新的計算技術時代。
從最早的帕斯卡爾齒輪機到今天最先進的電子計算機,其計算方式都是一種物理性質的符號變換,具體是由“加”和“減”這種基本動作構成的。然而,目前的DNA計算則有了本質性的變化。計算不再是一種物理性質的符號變換,而是一種化學性質的符號變換,即不再是物理性質的“加”、“減”操作而是化學性質的切割和粘貼、插入和刪除。這種計算方式的變革是前所未有的。具有劃時代的意義。
我們知道,DNA分子是一條雙螺旋的長鏈,上麵布滿了“珍珠”即核苷酸,其上擁有四種堿基,分別為:腺嘌呤(A)、鳥嘌呤(G)、胞嘧啶(C)和胸腺嘧啶(T)。DNA分子通過這些核苷酸的不同排列,能夠表達出生物體各種細胞擁有的大量信息。數學家、生物學家、化學家以及計算機專家從中得到啟迪。他們利用DNA能夠編碼信息的特點,先合成具有特定序列的DNA分子,使它們代表要求解的問題,然後通過生物酶的作用(相當於加減乘除運算),使他們相互反應,形成各種組合,最後過濾掉非正確的組合而得到的編碼分子序列就是正確答案。
這些都是我今後要努力的方向。謝謝您的提示。
- [1]wangbin6087
- 博主的觀點可以為計算主義提供辯護,並可以從數學的角度考察“演化”和“計算”這兩個概念的同質性,從而使人們對“計算”的意義有更本質的理解。
- 博主回複(2017-7-23 20:05):謝謝您的評論和推薦。從數學或計量(計算)分析研究生命的本質及其發展規律不是我的獨創,但我確實是在前人研究的基礎上進行了突破與發展。馬克思講,"一門學科隻有當它應用數學時候,這門學科才能算是成熟的科學“。生命科學現在還不能算是真正意義上的科學,因為生命科學在計量分析這方麵做的還遠遠不夠,與物理學等等其他學科相比還存在著嚴重不足。我的這篇論文隻是想在這方麵做出點學術貢獻。不足之處還請多多批評指正。
