付巧妹的博士導師是2022年諾貝爾獎得主斯萬特·帕博。
該文發表時間:2023年6月。中文媒體幾乎沒有報道。
東亞人進入亞洲的事實已無法掩蓋。
人類學學報, 2023, 42(03): 412-421 doi: 10.16359/j.1000-3193/AAS.2023.0010
綜述
古基因組揭示史前歐亞大陸現代人複雜遺傳曆史
張明 ,1,2,3, 平婉菁2,3, 付巧妹,2,3
,1,2,3, 平婉菁2,3, 付巧妹,2,3
1.中國-中亞人類與環境“一帶一路”聯合實驗室, 文化遺產研究與保護技術教育部重點實驗室, 西北大學文化遺產學院, 西安710127
2.中國科學院古脊椎動物與古人類研究所, 中國科學院脊椎動物演化與人類起源重點實驗室, 北京100044
3.中國科學院生物演化與環境卓越創新中心, 北京100044
4.裏士滿大學生物係, 裏士滿VA 23173
Ancient genomes reveal the complex genetic history of Prehistoric Eurasian modern humans
ZHANG Ming,1,2,3, PING Wanjing2,3, YANG Melinda Anna2,4, FU Qiaomei,2,3
1. China-Central Asia “the Belt and Road” Joint Laboratory on Human and Environment Research, Key Laboratory of Cultural Heritage Research and Conservation, School of Culture Heritage, Northwest University, Xi’an 710127
2. Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy of Sciences, Beijing 100044
3. CAS Center for Excellence in Life and Paleoenvironment, Beijing 100044
4. Department of Biology, University of Richmond, Richmond, VA 23173, USA

古DNA提取與測序技術的發展,讓科學家們能夠利用史前現代人樣本直接開展古基因組研究。古基因組研究發現,在約4萬年前,歐亞大陸上至少存在著6個獨立的現代人群體,其中3個群體並未對現今人群貢獻基因。在距今4萬年至末次盛冰期結束,歐亞大陸至少有5個具有代表性的現代人群體。末次盛冰期結束後的部分現代人群體與現今人群擁有更近的遺傳關係,而部分群體則未對現今人群貢獻基因。本文基於末次盛冰期前(45~19 kaBP)與末次盛冰期後(19~10 kaBP)兩個重要時間段的歐亞大陸史前現代人的基因組研究,梳理歐亞現代人在時間與空間上的發展脈絡,重點探究此前研究相對滯後的歐亞大陸東部地區。
關鍵詞: 歐亞大陸; 現代人; 古DNA; 人群結構; 遺傳曆史
Significant shifts in human populations occurred several times throughout history, as populations dispersed throughout Eurasia about 50 kaBP. During the Last Glacial Maximum (LGM), global temperatures dropped sharply causing environmental deterioration and population turnover in areas. After the LGM, populations increased as the natural environment stabilized and gradually developed into today’s populations. With advancements in ancient DNA extraction and sequencing technology, it is increasingly possible to directly retrieve genome-wide data from prehistoric modern human remains. The rapid emergence of new ancient genomes provides an entirely new direction for studying modern human population structure and evolutionary history. This research on Eurasian populations spanning 45~19 kaBP (pre-LGM) and 19~10 kaBP (post-LGM) summarizes the movement and interaction of prehistoric modern human populations, focusing especially on prehistoric East Eurasia, a region that has been less well-studied genetically. Of at least six distinct populations in Eurasia, three did not contribute substantial ancestry to present-day populations: Ust’-Ishim (≈45 kaBP) from northwestern Siberia; Oase 1 (≈40 kaBP) from Romania; and Zlatý k?ň (over 45 kaBP) from Czechia. One population represented by three individuals (4.6~4.3 kaBP, from Bacho Kiro Cave, Bulgaria) seemed to contribute at least a partial genetic component to later some Eurasian populations. One population represented by Tianyuan man (≈40 kaBP, from East Asia) was shown to be more similar to present-day East Asians and Native Americans than to present-day or ancient Europeans. One population represented by Kostenki 14 (≈36 kaBP, from western Siberia) and Goyet Q116-1 (≈35 kaBP, from Belgium) was more closely related to Europeans than to other Eurasians. This work also summarized five representative populations after 40 kaBP and before the end of the LGM. In East Eurasia after the LGM (or since 14 kaBP), population histories played out very differently. For instance, high genetic continuity is observed in the Amur region in the last 14 kaBP, while in the Guangxi region of southern China, an ancient population that lived 10.5 kaBP carried ancestry not represented in any present-day humans. To conclude, comparison of genome-wide ancient DNA from multiple prehistoric humans have illustrated a complex genetic history of prehistoric Eurasian modern humans. In the future, additional ancient genomes will provide more evidence and details to illuminate the complex genetic history of modern humans.
Keywords: Eurasia; modern human; Ancient DNA; Population structure; Genetic history
本文引用格式
張明, 平婉菁, 付巧妹. 古基因組揭示史前歐亞大陸現代人複雜遺傳曆史[J]. 人類學學報, 2023, 42(03): 412-421 doi:10.16359/j.1000-3193/AAS.2023.0010
ZHANG Ming, PING Wanjing, YANG Melinda Anna, FU Qiaomei. Ancient genomes reveal the complex genetic history of Prehistoric Eurasian modern humans[J]. Acta Anthropologica Sinica, 2023, 42(03): 412-421 doi:10.16359/j.1000-3193/AAS.2023.0010
1 引言
古基因組研究表明,歐亞大陸在史前時期存在多個現代人群體,且這些人群的結構在50 kaBP以來發生了幾次重要改變。現代人快速且成功地擴散至整個歐亞大陸乃至於美洲[1⇓-3],可能與這一時期地球氣候的幾次冷暖更替事件相關[4]。末次盛冰期(約26.5 kaBP開始,一直持續到約19 kaBP)時,全球氣溫驟降,自然環境極度惡化,一些地區的現代人麵臨著嚴峻的生存挑戰,有些地區的人群結構可能因此而發生顛覆性的改變,造成冰期後再遷入的人群與之前的人群不存在直接的基因聯係[5]。末次盛冰期之後,地球環境進入相對穩定並且適宜人類生存的時期,各地的現代人群體不斷發展、遷徙、融合,逐漸形成了現今現代人的群體結構。
近年來,古DNA實驗技術與研究方法的高速發展[6⇓⇓-9],極大地推動了人類學的研究。通過古DNA技術直接獲取史前材料中的遺傳信息,結合材料發現的地點和測年信息,研究者們得以在一個更大的時空框架下開展綜合分析。相比於傳統的化石形態學研究,古DNA研究對於材料完整度的要求相對較低,並且能夠提供更為客觀的數據信息。此外,由於每一個個體的古基因組數據都能夠在很大程度上反映群體的遺傳特征,所以單個個體的古基因組數據能夠代表整個群體進行分析,這對於隻發現零星人骨遺存的遺址來說,具有更加重要的意義。自2010年,4 kaBP的愛斯基摩人、尼安德特人及丹尼索瓦人的基因組數據發布以來[6,10,11],已經有超過6000例人類古基因組數據發布[12],相關古基因組研究發現和結論已經證實、推翻或修正了古人類學領域許多已有的觀點和假說,並提出了許多新的科學認知。如,證實除尼安德特人外還存在另外一種滅絕古人類──丹尼索瓦人[11],發現滅絕古人類(尼安德特人和丹尼索瓦人)與現代人有過基因交流[6,11],推翻了此前認為的現代人與尼安德特人自分離後不再有基因交流的觀點;還修正了此前歐亞大陸東部地區的“兩層假說”,發現新石器時代早期中國南方福建等沿海地區古人群,實質上並無明顯屬於“第一層”人群的遺傳信息[13]。2017年以前,已發表的古基因組數據主要集中於歐亞大陸西部,科學家們對歐洲、北亞等區域的人群演化問題已展開比較詳盡的研究。2017年以來,隨著科學家們把關注的重心移向歐亞大陸東部地區,歐亞大陸東部人類古基因組數據開始持續發布,並湧現出了諸多重要的成果,使這一區域人類古基因組研究相對落後的局麵已開始逐漸改變。本文重點關注歐亞大陸東部地區史前現代人相關的重要古基因組研究成果,希望為讀者梳理、總結歐亞現代人,尤其是歐亞東部現代人在時間與空間上的發展脈絡。
2 遺傳曆史
圖1
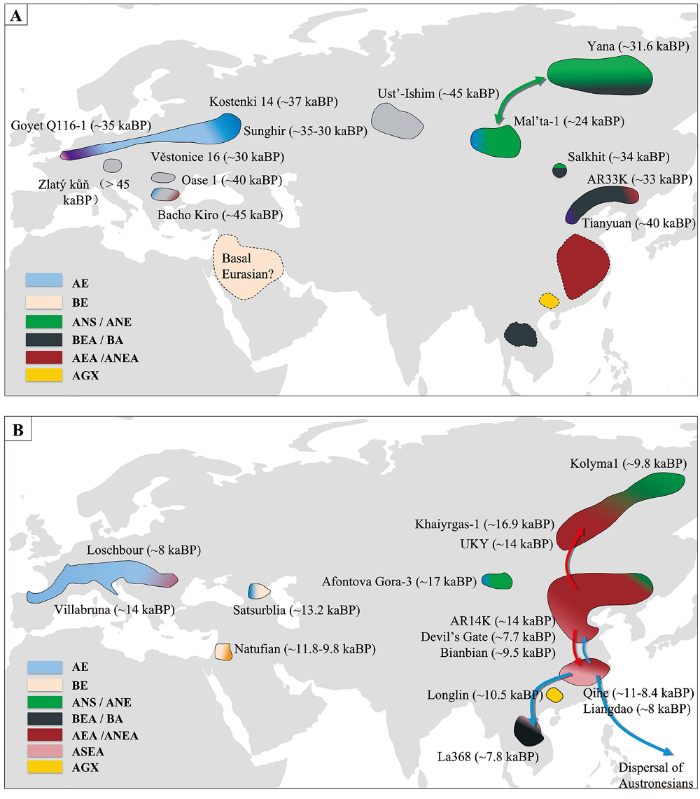
圖1 45~10 kaBP歐亞大陸人群示意圖
A)階段1 Period I(45~19 kaBP);B)階段2 Period II(19~10 kaBP):AE,以Kostenki 14為代表的古歐洲人群相關的遺傳成分ancestry related to Ancient Europeans (represent by Kostenki 14);BE,與未經取樣的古歐亞人群相關且在現今歐洲人群、部分古代和現今中東人群中可找到的遺傳成分ancestry related to an unsampled population known as Basal Eurasians and found in small numbers in ancient and present-day populations of the Near East and in present-day Europeans;ANS,以Yana為代表的古西伯利亞北部人群相關的遺傳成分ancestry related to ancient North Siberians (represent by Yana); ANE, ancestry related to ancient North Eurasians (represent by Mal’ta-1 and Afontova Gora-3);ANE,以Mal’ta-1和Afontova Gora-3為代表的古北歐亞人群相關的遺傳成分;BEA,以田園洞人和AR33K為代表的基礎東亞人相關的遺傳成分;BA,以La368為代表的古代東南亞狩獵采集人群相關祖源成分;AEA,東亞古北方人群和東亞古南方人群分離前的亞洲人群;ANEA,以AR14K、扁扁洞、Devil’s Gate等個體為代表的東亞古北方人群;ASEA,以奇和洞、亮島個體為代表的東亞古南方人群;AGX,以隆林個體為代表的古廣西人群。圖1A中的紫色代表田園洞人和Goyet Q116-1個體相聯係的成分。虛線區域是指該遺傳成分源自根據遺傳數據所推測的人群狀態,但未發現這一時期的古代樣本。各顏色大致顯示為某代表區域之間或之內的遺傳成分組合,且顏色梯度顯示為不同人群之間可能存在的遺傳聯係(如基因流)。
Fig.1 Schematic of Populations in Eurasia from 45 to 10 kaBP
AE, ; BE, ; ANS, ; BEA, ancestry related to Basal East Asians (represent by Tianyuan and AR33K); BA, ancestry related to Basal Asians (represented by La368); AEA, ancestry related to Ancient East Asians; ANEA, ancestry related to Ancient Northern East Asians (represented by AR14K, Bianbian, Devil’s Gate, etc); ASEA, ancestry related to Ancient Southern East Asians (represented by Qihe and Liangdao); AGX, ancestry related to Ancient Guangxi population (represented by Longlin). The purple color in Fig.1A shows the connection between Tianyuan and Goyet Q116-1. Broken lines indicate no ancient genetic samples have been found for a population with the inferred ancestry. Colors loosely indicate genetic groupings between or within a region, with color gradients showing the connections (i.e., gene flow) that may exist between different ancient populations.
2.1 階段1(45~19 kaBP)
歐亞大陸史前現代人的古基因組研究表明,在約40 kaBP前,歐亞大陸至少存在6個不同的現代人群體(圖1),分別為:
同樣,Oase 1個體與14 kaBP及更早期的古歐洲人群、現今東亞人群和現今美洲原住民人群均擁有相似數量的等位基因,即擁有相近的親緣關係。這一結果表明Oase 1個體所代表人群在歐洲無延續至今的直接後裔,是一種對現今歐亞人群無實質遺傳貢獻的獨特人群;而且由於攜有6%~9%的尼安德特人遺傳成分,Oase 1個體可能代表了與尼安德特人具有密切互動關係的一支歐洲早期現代人群體[7]。
Zlatý k?ň個體所代表的人群,同樣未給之後的歐洲人和亞洲人貢獻基因成分。與同時期的其他現代人個體相似,Zlatý k?ň個體攜有約3%的尼安德特人成分,但其含有尼安德特人基因片段的平均長度長於Ust-Ishim個體,這說明發生的基因重組次數少,前者的尼安德特人成分混入時間短於後者。Zlatý k?ň個體所代表的人群也是最早定居歐亞大陸的人群之一[18]。
2)以發現於保加利亞的46~43 kaBP的Bacho Kiro洞的3個個體為代表的古人群。這一人群相比於晚些時間出現在歐亞大陸西部地區的人群,與東亞和美洲的現代和古代人群關係更近。這表明他們屬於一個早前遷入歐洲、但此前沒有發現基因證據的群體,而且至少有部分與這一群體相關的基因成分延續到了後期的歐亞大陸人群中[19]。
3)以發現於中國北京約40 kaBP的田園洞人[20⇓-22]為代表的古人群,可稱之為“古東亞人”(Basal East Asians, BEA)。這一人群廣泛分布在歐亞大陸東部地區,與稍晚一些生存在黑龍江流域的AR33K個體(約33.6 kaBP)屬於同一人群。研究發現,田園洞人較之現今歐洲人群和古歐洲人群來說,與現今東亞人群和美洲原住民人群有更近的親緣關係[22]。相較於其他美洲原住民,某些南美洲原住民人群,特別是蘇魯人(Suruí)和卡利吉亞納人(Karitiana),顯示與大洋洲人、安達曼昂格人和田園洞人均有遺傳聯係,這表明美洲原住民人群的祖先們可能分批進入美洲,因而亞洲人群對其有不同程度的遺傳貢獻[22,23]。這些聯係也表明,田園洞人的遺傳成分至少存在於歐亞東部人群的亞群之中,且以某種形式一直延續到歐洲殖民者進入美洲之前。
4)以發現於西伯利亞西部約36 kaBP的Kostenki 14個體和發現於比利時約35 kaBP的Goyet Q116-1個體[24,25]為代表的古人群,可稱之為“古歐洲人”(Ancient Europeans, AE)。“古歐洲人”較之於其他歐亞人群而言,與現今歐洲人群的關係更為相近[24,25],這表明某些與現今歐洲人群有遺傳聯係的現代人群體至少在35 kaBP已經廣泛分布於歐亞大陸西部[24](圖1)。發現於俄羅斯約34 kaBP的Sunghir個體與Kostenki 14個體及約34~26 kaBP的捷克V?stonice 16個體均有遺傳聯係,這意味著Kostenki 14所代表人群,至少有一部分被與V?stonice 16有親緣關係的人群所替代[26]。
綜合田園洞人、Kostenki 14、Goyet Q116-1等個體的古基因組研究可以發現,在約40 kaBP,遺傳學意義上的歐洲人群已在歐洲(比利時)出現,遺傳學意義上的亞洲人群也已在亞洲(中國)出現[22]。這也表明歐亞人群的分化(即歐洲人群和亞洲人群的分離)很可能早於40 kaBP,這與依據遺傳突變率所推導的這兩個人群的分歧時間(40~20 kaBP前至80~40 kaBP)一致[27⇓-29]。然而,雖然田園洞人與Goyet Q116-1個體在地理上相隔萬裏,但他們之間卻存在著遺傳聯係。此外,他們分別與現今亞洲人群和歐洲人群具有更近的遺傳聯係,但是他們所共有的等位基因數量比早期亞洲人群和歐洲人群之間以簡單分離模式所預期的多[22],這種現象產生的原因可能與亞洲人群和歐洲人群在分離之後仍發生過基因交流相關[30⇓-32]。考慮到Goyet Q116-1個體和田園洞人這兩個個體生存時間之早和地理距離之遠,Yang等認為當時的東亞人可能並未直接與歐洲人發生基因交流,而可能是跟某一個與這兩個個體代表人群均相關的未知人群都發生過基因交流,而這個未知人群可能是從尚未分化的古歐亞人群中的某一亞群演化而來[22]。近期的一項研究發現,保加利亞46~43 kaBP的Bacho Kiro古人群,將Goyet Q116-1個體和田園洞人代表的兩個人群聯係了起來[19]。
距今4萬年以來至末次盛冰期前的現代人群體中有些群體獨立存在,還有一些群體與距今4萬年以前的古人群相聯係,比較具有代表性的有以下5個群體:
2)以發現於蒙古國東北部約34 kaBP的Salkhit個體為代表的古人群。Salkhit個體屬於歐亞大陸東西部人群分離後形成的人群,而且有歐亞大陸西部人群的遺傳成分。Salkhit個體攜帶有約75%的與田園洞人相關的祖源成分(BEA相關祖源成分),以及約25%的與Yana人群相關的祖源成分(ANS相關祖源成分)[35]。
3)以發現於中國黑龍江流域約33.6 kaBP的AR33K個體為代表的古人群。AR33K個體與田園洞人有最近的親緣關係,但是AR33K個體與Goyet Q116-1個體的關係並非像田園洞人與Goyet Q116-1個體一樣相近。而且,相比於其他美洲原住民人群,AR33K個體也沒有與南美洲的蘇魯人(Surui)擁有更近的關係。另外,AR33K個體中也沒有發現與Yana人群相關的祖源成分。這表明東亞北部地區可能在這一時期已經形成複雜的人群結構。田園洞人、AR33K個體及Salkhit個體的古基因組研究共同表明,末次盛冰期前,與田園洞人相關的人群在東亞北部地區廣泛分布(分布範圍至少覆蓋田園洞人代表的華北平原、AR33K代表的中國東北地區和Salkhit代表的蒙古國東北部地區),該人群在蒙古國與Yana相關人群產生了基因混合,而這一時期的黑龍江流域的人群則保持獨立特征[5]。
5)以現有遺傳數據推測出來的一個未知人群,可稱之為古歐亞人群(Basal Eurasians,BE),出現在歐洲人群和亞洲人群分離之前[36]。現今歐洲人群攜有這種古歐亞人群的遺傳成分,而這種遺傳成分的獲得主要發生在農業文明進入歐洲以後,因而相較於農業文明以前的古歐洲人群(即14 kaBP及更早期的不攜有古歐亞人群(BE)遺傳成分的古歐洲人群)而言,現今歐洲人群總體上與其他現代人群的關係較遠。Lazaridis等[37]研究發現,古歐亞人群(BE)遺傳成分在約34~14 kaBP前的近東人中存在較高比例,在約13 kaBP前高加索地區的狩獵采集人群中也開始發現古歐亞人群(BE)的遺傳成分。另外,該人群顯示同時具有歐洲人群和近東黎凡特地區的納圖夫人群(Natufians)的遺傳特征。歐洲西部的狩獵采集人群與歐洲東部的非洲以外人群(non-African)至少在40 kaBP前就已經分離,而古歐亞人群的分歧時間更早,可能在非洲以外人群與滅絕古人類發生基因混合之前就已經分離[36,37],因此可能不具有或僅具有極低含量的尼安德特人成分[37]。古歐亞人群形成的時間相對較早,存在的時間可能比較長,且持續到更晚的時期,對歐洲和近東地區的現代人產生重要的遺傳影響。現今歐洲人群所攜有的尼安德特人成分低於現今歐亞地區的其他現代人[9,37⇓-39],很可能是因為古歐亞人群後期進入歐洲,導致歐洲人的尼安德特人成分被稀釋。
2.2 階段2 (19~10 kaBP)
末次盛冰期結束後,歐亞大陸各地區的現代人進入快速發展期。這一時期有些現代人群體與現今現代人群體之間存在一定的遺傳連續性,也正是這一時期不同地域古人群的擴增、遷徙、融合等活動,逐漸奠定了現今現代人的遺傳結構和分布格局。
在歐亞大陸東北部地區,末次盛冰期前後的人群不連續,但末次盛冰期結束後的人群則存在連續性。發現於中國黑龍江流域19 kaBP的AR19K個體相比於末次盛冰期前的古東亞人(如田園洞人和AR33K)而言,與末次盛冰期後至全新世的人群具有更大的遺傳相似性,這表明在末次盛冰期即將結束時,東亞東北部的人群結構已經產生了顛覆性改變[5],這一情況不同於貝加爾湖區域人群在末次盛冰期前後的連續性[24,34]。相比於東亞古南方人群(Ancient Southern East Asians, ASEA;以中國福建約8.4 kaBP的奇和洞個體和台灣海峽約8.3 kaBP的亮島個體為代表),AR19K個體與東亞古北方人群(Ancient Northern East Asians, ANEA;以中國山東9.5~7.7 kaBP的個體為代表)遺傳關係更近。這表明在末次盛冰期結束時,東亞南北方古人群的遺傳差異已經形成,而AR19K個體所代表的人群也被認為是最早期的東亞古北方人群(ANEA)[5]。這一人群後期以黑龍江流域的一係列時間連續的個體(AR14K等)、俄羅斯濱海地區(鬼門洞Devil’s Gate)約7.7 kaBP的個體、中國山東9.5~7.7 kaBP的個體(如扁扁洞Bianbian)、貝加爾湖區域約7.1~6.3 kaBP的個體作為代表[2,5,13,40]。自14 kaBP以來,這一人群在中國黑龍江流域具有遺傳連續性,現今這一區域的一些少數民族仍然攜有這些人群延續下來的遺傳成分[40]。
這一時期分布在西伯利亞的古人群顯示出不同祖源成分的混合,他們對後期出現的人群產生了重要影響。該人群被稱之為古西伯利亞人群(Ancient Paleo-Siberians, APS),以發現於俄羅斯貝加爾湖地區約14 kaBP的UKY個體、遠東地區約9.8 kaBP的Kolyma1個體為代表。這一人群以亞洲祖源成分為主,但混有古西伯利亞北部人群(ANS)相關的祖源成分,且Kolyma1代表著在美洲之外與美洲原住民人關係最近的人群[2,42]。在俄羅斯西伯利亞發現的約17 kaBP的Khaiyrgas-1個體所代表的群體與古西伯利亞人群(APS)具有十分相近的遺傳關係[41],是該人群的早期代表之一。研究表明古西伯利亞人群(APS)的東亞祖源成分的最近來源便是14 kaBP以來的東亞古北方人群(ANEA)[5],這表明古西伯利亞人群(APS)可能與東亞古北方人群(ANEA)發生過基因混合。而古西伯利亞人群(APS)相關祖源成分也在山東博山(Boshan)、鬼門洞(Devil’s Gate)、貝加爾湖個體中存在,這表明至少有部分遺傳成分從古西伯利亞人群(APS)混入到東亞古北方人群(ANEA)中,說明至少在8 kaBP前,古西伯利亞人群(APS)已經與東亞古北方人群(ANEA)有所聯係[5,13,43]。
除與現今現代人有遺傳聯係的古人群外,東亞南部還發現了未對現今現代人產生遺傳影響的史前現代人群體。發現於廣西隆林(Longlin)約10.5 kaBP的個體所代表的古廣西人群(Ancient Guangxi population, AGX),與此前發布的中國南方沿海的東亞古南方人群(ASEA,以此前福建個體為代表)及東南亞狩獵采集人群相關祖源人群(基礎亞洲人Basal Asians, BA;以發現於老撾約8 kaBP的La368個體和馬來西亞約4.3 kaBP的Ma911個體為代表的Hòabìnhian狩獵采集人群)[44]不同,屬於此前未知的東亞現代人群體,對現今現代人未產生遺傳影響。這一人群與東亞南北方祖源人群(ASEA和ANEA)具有很深分歧,並且至少持續到6 kaBP。而約9~6 kaBP的廣西人群混合有當地祖源成分(AGX)、東亞古南方祖源成分(ASEA)及東南亞狩獵采集人群相關祖源成分(BA)[44],這表明了農業在這一區域興起之前,人群已經有了廣泛的混合,也說明在東亞和東南亞存在的3個獨立祖源人群在史前時期互動頻繁。
這一時期的歐亞大陸西部人群相比於更古老的歐洲人群,顯示出與現今亞洲人群和近東人群更近的親緣關係。約14 kaBP以來的歐洲中西部個體與一些跨越舊石器和新石器時代的文化個體(如意大利的Villabruna個體)密切相關,且顯示出與現今近東人群和東亞人群有遺傳聯係[22,24,34]。高加索地區約13~10 kaBP的個體(如格魯吉亞的Satsurblia和Kotias)與歐亞西部個體有較近的親緣關係,且含有古歐亞人群(BE)的遺傳成分。古歐亞人群(BE)的遺傳成分在現今歐洲人群和近東人群中均有存在[24,36,37],尤其在近東人群中的含量最高,推算在舊石器時代晚期約12~9.8 kaBP的黎凡特地區,納圖夫人(Natufian)個體中古歐亞人群(BE)遺傳成分的含量高達44%;在中石器時代文化約9.1~8.6 kaBP的伊朗個體(如伊朗的Hotu個體)中古歐亞人群(BE)遺傳成分含量高達66%[37]。
除以上列舉的主要史前現代人群體外,科學家們對全新世以來的更多材料進行了古基因組學研究,發現不同人群之間產生了多次交互影響的事件,對世界現代人群體的分布起到重要的影響。如:在貝加爾湖區域,新石器早期古北歐亞人(ANE)祖源人群,被與現今東亞北部人群相關的祖源人群大量替代[45]。進入全新世之後,東亞南北方人群頻繁互動,但整體而言主要有三次大規模的南向基因流動,包括:1)東亞古北方人群相關的遺傳成分在東亞南方人群中有所增加;2)具有東亞遺傳成分的人群向東南亞擴散,並與基礎亞洲人(BA)混合;3)東亞古南方人群祖源成分,通過南島語係人群向東南亞島嶼和太平洋島嶼擴散[13,43]。人群的互動為現今的東亞大陸人群遺傳格局的形成奠定了基礎。
3 展望
史前現代人群體的古基因組研究揭示了歐亞大陸豐富的人類曆史。對這一時期的現代人展開的古基因組研究,已揭示出歐亞大陸人群豐富多彩的遺傳演化曆史,極大地提升了我們對史前人群的更替和遷徙曆史等相關問題的理解和認識[48]。近年來,歐亞大陸東部的人類古基因組研究快速發展,尤其一些東亞南方樣本基因組信息的成功獲取,為我們發現和了解更多史前人類群體的遺傳曆史提供了重要支撐[13,47]。雖然歐亞大陸史前現代人基因混合和人群結構相關的許多問題已得到了答案,但隨著研究的深入,新的科學問題也在不斷出現,例如,滅絕古人類對不同區域的史前現代人和現今現代人群體的基因貢獻程度及影響如何?古基因組是否能發現更多獨立的史前現代人群體?史前現代人如何發展、演變、交流,並逐漸形成後期人群及現今人群格局的?
隨著古DNA實驗技術的日益成熟和分析方法的不斷完善,研究者們也能夠擴展除歐亞大陸以外的材料以進行研究,有助於更全麵揭示早期現代人的群體結構,以及滅絕人類在現代人演化曆史中所扮演角色等相關問題。近年來,科學家們開展了許多關於新石器時代以來現代人群的古基因組研究,尤其是東亞[5,13,47,49,50]、歐洲[8,35,36]和美洲[51]人群。古基因組研究團隊在長時間尺度下開展人群遺傳演化分析時需要盡量結合多個學科,從不同角度進行探討,才能夠更好地還原真實的人群演化圖景。
 選擇“Disable on www.wenxuecity.com”
選擇“Disable on www.wenxuecity.com”
 選擇“don't run on pages on this domain”
選擇“don't run on pages on this domain”