馬可安博客
馬可安在全球首先提出中國華北嚴重霧霾的核霧染成因理論,成為唯一被當局組織全麵討伐圍剿的霧霾成因理論,我利用本博客進行進一步的學術探討。



正文
從科學和邏輯的角度解釋為什麽武漢冠狀病毒是人造的
?自從疫情發生以來,關於武漢冠狀病毒的源頭有非常多的爭議。可以這麽說,它的兩種可能來源(自然進化或人造)似乎都有一定依據。對此全世界理應充分的分析和討論。在社交媒體上,兩種說法基本勢均力敵。可是在科研論文或其他文章中,支持病毒來源於自然並譴責所謂“陰謀論”的占絕大多數。而支持另一邊的,就是認為病毒來源於人造的,其科學論文或其他文章則幾乎沒有。在這篇文章裏,我將用科學證據和邏輯推理來證明,這個武漢冠狀病毒絕不是來源於自然。
我的分析會建立在確實可靠的證據之上。同時,我會主動排除不可信的,甚至是可疑的所謂證據。因為有些“證據”很有可能是中共主動摻進來,已達到混淆視聽和掩蓋真相之目的。鑒於此,我在分析中將扮演一個集科學家,偵探,和法官三重身份為一體的一個角色。
人們為什麽覺得武漢冠狀病毒的來源可疑
這種想法很大程度上源於對病毒序列的比較和分析。
一般有兩種比較序列的方式,一個是比較基因序列,一個是比較蛋白質序列。對於病毒來說,這兩種比較基本等同。這是因為病毒的基因序列基本全部被翻譯成蛋白(病毒一般是一次性把自己的整個基因組翻譯成一個長肽鏈,然後再用蛋白酶把它切成各個單獨的蛋白)。所以我們在這兒就隻用蛋白序列來比較幾個相關的病毒。
通過這樣的序列比較,很明顯可以看出武漢冠狀病毒和2003年的SARS冠狀病毒有86%左右的一致性。這個程度的一致性基本就是說,兩個病毒是同一類,但從進化角度來說,武漢冠狀病毒肯定不是從SARS進化來的。這一點也是病毒領域裏大家公認的。
值得注意的是,武漢冠狀病毒與兩個蝙蝠冠狀病毒有極其詭異的相似性。這兩個蝙蝠病毒(ZC45和ZXC21)統稱為舟山蝙蝠病毒,因為都是從浙江舟山采集到的。從全序列的比較來看,舟山蝙蝠病毒和武漢冠狀病毒一致性達到了95%。而對於序列裏的絕大多數蛋白來說,這個一致性是普遍的,有的甚至更高,比如E蛋白的一致性是100%。Nucleocapsid蛋白是94%,membrane蛋白(膜蛋白)是98.6%,S2蛋白(spike蛋白的後半部分)是95%。然而,詭異的是,S1蛋白,也就是spike蛋白的前半部分,非常與眾不同。在這裏,兩個病毒序列的一致性突然降到了69%。這種一致性的分布(所有其他部位95%,而僅一個特定蛋白69%)從遺傳進化的角度來講是極其詭異的。
說它詭異是因為自然進化絕大多數情況下是通過隨機突變的方式進行的。那麽假如僅僅是隨機突變的話,祖先和後代之間序列上的差異是應該基本均勻分布的。或者說各個蛋白之間的一致性應該都差不多,不應該出現一個蛋白如此與眾不同的局麵。
那麽有沒有其他的進化方式會導致這樣奇特的蛋白一致性的分布呢?是的,有一種方式可以做到,就是基因重組。我會在之後的段落裏詳細分析為什麽天然的基因重組也不可能使舟山蝙蝠病毒或其近親進化成為武漢冠狀病毒。
現在,讓我們把目光聚焦在這個與眾不同的S1蛋白身上。
我的分析會建立在確實可靠的證據之上。同時,我會主動排除不可信的,甚至是可疑的所謂證據。因為有些“證據”很有可能是中共主動摻進來,已達到混淆視聽和掩蓋真相之目的。鑒於此,我在分析中將扮演一個集科學家,偵探,和法官三重身份為一體的一個角色。
人們為什麽覺得武漢冠狀病毒的來源可疑
這種想法很大程度上源於對病毒序列的比較和分析。
一般有兩種比較序列的方式,一個是比較基因序列,一個是比較蛋白質序列。對於病毒來說,這兩種比較基本等同。這是因為病毒的基因序列基本全部被翻譯成蛋白(病毒一般是一次性把自己的整個基因組翻譯成一個長肽鏈,然後再用蛋白酶把它切成各個單獨的蛋白)。所以我們在這兒就隻用蛋白序列來比較幾個相關的病毒。
通過這樣的序列比較,很明顯可以看出武漢冠狀病毒和2003年的SARS冠狀病毒有86%左右的一致性。這個程度的一致性基本就是說,兩個病毒是同一類,但從進化角度來說,武漢冠狀病毒肯定不是從SARS進化來的。這一點也是病毒領域裏大家公認的。
值得注意的是,武漢冠狀病毒與兩個蝙蝠冠狀病毒有極其詭異的相似性。這兩個蝙蝠病毒(ZC45和ZXC21)統稱為舟山蝙蝠病毒,因為都是從浙江舟山采集到的。從全序列的比較來看,舟山蝙蝠病毒和武漢冠狀病毒一致性達到了95%。而對於序列裏的絕大多數蛋白來說,這個一致性是普遍的,有的甚至更高,比如E蛋白的一致性是100%。Nucleocapsid蛋白是94%,membrane蛋白(膜蛋白)是98.6%,S2蛋白(spike蛋白的後半部分)是95%。然而,詭異的是,S1蛋白,也就是spike蛋白的前半部分,非常與眾不同。在這裏,兩個病毒序列的一致性突然降到了69%。這種一致性的分布(所有其他部位95%,而僅一個特定蛋白69%)從遺傳進化的角度來講是極其詭異的。
說它詭異是因為自然進化絕大多數情況下是通過隨機突變的方式進行的。那麽假如僅僅是隨機突變的話,祖先和後代之間序列上的差異是應該基本均勻分布的。或者說各個蛋白之間的一致性應該都差不多,不應該出現一個蛋白如此與眾不同的局麵。
那麽有沒有其他的進化方式會導致這樣奇特的蛋白一致性的分布呢?是的,有一種方式可以做到,就是基因重組。我會在之後的段落裏詳細分析為什麽天然的基因重組也不可能使舟山蝙蝠病毒或其近親進化成為武漢冠狀病毒。
現在,讓我們把目光聚焦在這個與眾不同的S1蛋白身上。
?圖1. 冠狀病毒的模擬圖。紅色的就是spike蛋白。圖片來源於美國疾病控製中心(CDC)
Spike蛋白就是裝飾在病毒表麵的紅色的凸起(圖1)。冠狀病毒的名字就是從它而來,因為它使病毒在電鏡下看起來就像皇冠。從功能角度來說,spike蛋白非同小可,它是冠狀病毒的“鑰匙”,可以打開我們(宿主)細胞的“鎖”,從而進入並侵染細胞。
圖2顯示的是SARS的spike蛋白的結構。你可以把這樣的結構想象為給真人拍的照片,它就是這個蛋白的真實麵目。因為武漢冠狀病毒和SARS冠狀病毒的序列一致性也相當高,所以武漢冠狀病毒的spike蛋白在結構上應該和這個SARS的spike基本一致。最近剛剛發表的文章也印證了這一點(1)。
Spike蛋白就是裝飾在病毒表麵的紅色的凸起(圖1)。冠狀病毒的名字就是從它而來,因為它使病毒在電鏡下看起來就像皇冠。從功能角度來說,spike蛋白非同小可,它是冠狀病毒的“鑰匙”,可以打開我們(宿主)細胞的“鎖”,從而進入並侵染細胞。
圖2顯示的是SARS的spike蛋白的結構。你可以把這樣的結構想象為給真人拍的照片,它就是這個蛋白的真實麵目。因為武漢冠狀病毒和SARS冠狀病毒的序列一致性也相當高,所以武漢冠狀病毒的spike蛋白在結構上應該和這個SARS的spike基本一致。最近剛剛發表的文章也印證了這一點(1)。
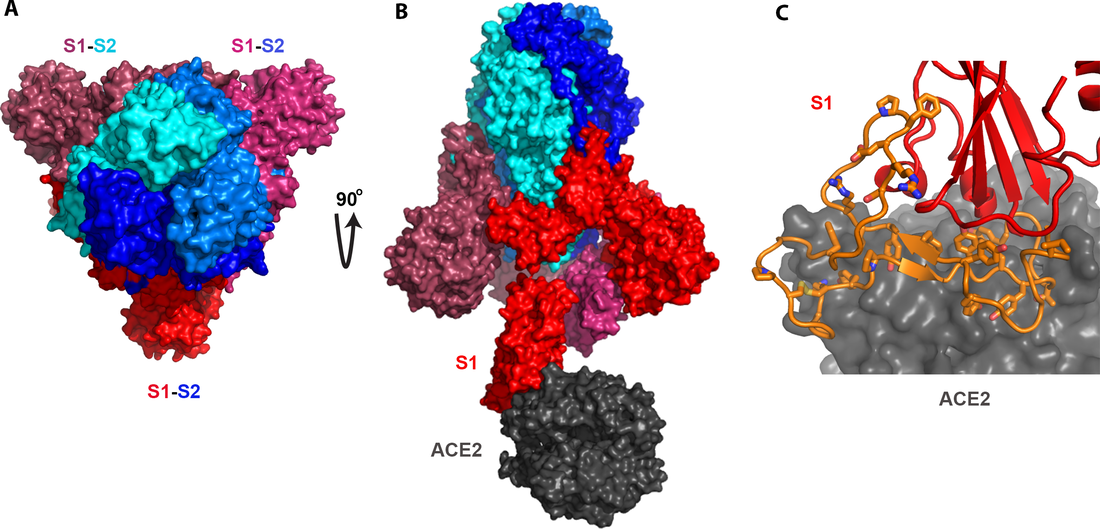
?圖2. SARS病毒spike蛋白的結構以及它如何結合人體細胞上的受體ACE2。此圖的製作是基於已發表的結構PDB ID: 6acj(2)。A)三個spike蛋白形成三聚體,每個spike蛋白都由兩個基本等長的部分組成,S1和S2。B)S2部分(藍色)負責三聚體的形成,而S1部分決定和細胞受體的結合。在SARS這裏,受體是人體細胞的ACE2。C)此圖進一步揭示S1和ACE2結合的具體細節。橙色部分就是S1中和ACE2結合最關鍵的一段肽鏈。此橙色部分囊括了所有和ACE2結合的細節。其中最關鍵的氨基酸以棍狀結構做了標識。此段橙色序列就是後文中提到的,如同被從SARS的spike蛋白中“複製”出來,然後“插入”舟山蝙蝠病毒的spike蛋白中去的關鍵肽段。這一“操作”直接可以導致一個新的,能感染人的病毒的產生。
如圖2所示,三個spike蛋白聚集到一起形成所謂的三聚體。這個三聚體的形成完全取決於S2,也就是結構中的泛藍色部分。這個S2可以認為是一個“鑰匙”中用手抓著的那個部分。它不進入“鎖”裏,但對“鑰匙”來說又不可或缺(此“鑰匙”必須形成三聚體)。
Spike蛋白的另半部分就是S1,即圖中泛紅色部分。S1就是和細胞的受體結合的最關鍵的部分。你可以把S1想象成一把“鑰匙”中真正進入“鎖”的那部分。它必須和“鎖”(細胞受體)的內部的精細結構嚴絲合縫才能把細胞的“門”打開。一個“鎖”能不能被一個特定的spike“鑰匙”打開完完全全取決於這個S1。換句話說,S1決定了一個冠狀病毒會感染哪個宿主,哪種細胞。
了解了這些之後,你應該能更深刻的體會到為什麽我之前用“非常奇怪”或“極其詭異”來形容武漢冠狀病毒和舟山蝙蝠病毒之間序列對照的結果。當所有其他的部分都幾乎一樣的情況下,唯獨這個決定了病毒侵染哪個宿主的S1發生了極其顯著的改變。
好,我們再放大看一下具體是S1中的哪一小部分決定了和宿主細胞受體的結合(圖2C)。在這裏,和SARS的spike結合的受體是人體的ACE2蛋白。S1中對此結合起關鍵作用的這段肽鏈被標記為橙色,其中對結合ACE2起至關重要作用的氨基酸被標識成棍狀結構。這段橙色肽鏈囊括了所有和ACE2結合所需要的微觀結構。從下麵的分析中你會發現,這段本是SARS的spike蛋白所獨有的,並能充分決定spike蛋白和人體ACE2蛋白結合的肽段基本上被“複製”進了武漢冠狀病毒之中。
如圖2所示,三個spike蛋白聚集到一起形成所謂的三聚體。這個三聚體的形成完全取決於S2,也就是結構中的泛藍色部分。這個S2可以認為是一個“鑰匙”中用手抓著的那個部分。它不進入“鎖”裏,但對“鑰匙”來說又不可或缺(此“鑰匙”必須形成三聚體)。
Spike蛋白的另半部分就是S1,即圖中泛紅色部分。S1就是和細胞的受體結合的最關鍵的部分。你可以把S1想象成一把“鑰匙”中真正進入“鎖”的那部分。它必須和“鎖”(細胞受體)的內部的精細結構嚴絲合縫才能把細胞的“門”打開。一個“鎖”能不能被一個特定的spike“鑰匙”打開完完全全取決於這個S1。換句話說,S1決定了一個冠狀病毒會感染哪個宿主,哪種細胞。
了解了這些之後,你應該能更深刻的體會到為什麽我之前用“非常奇怪”或“極其詭異”來形容武漢冠狀病毒和舟山蝙蝠病毒之間序列對照的結果。當所有其他的部分都幾乎一樣的情況下,唯獨這個決定了病毒侵染哪個宿主的S1發生了極其顯著的改變。
好,我們再放大看一下具體是S1中的哪一小部分決定了和宿主細胞受體的結合(圖2C)。在這裏,和SARS的spike結合的受體是人體的ACE2蛋白。S1中對此結合起關鍵作用的這段肽鏈被標記為橙色,其中對結合ACE2起至關重要作用的氨基酸被標識成棍狀結構。這段橙色肽鏈囊括了所有和ACE2結合所需要的微觀結構。從下麵的分析中你會發現,這段本是SARS的spike蛋白所獨有的,並能充分決定spike蛋白和人體ACE2蛋白結合的肽段基本上被“複製”進了武漢冠狀病毒之中。
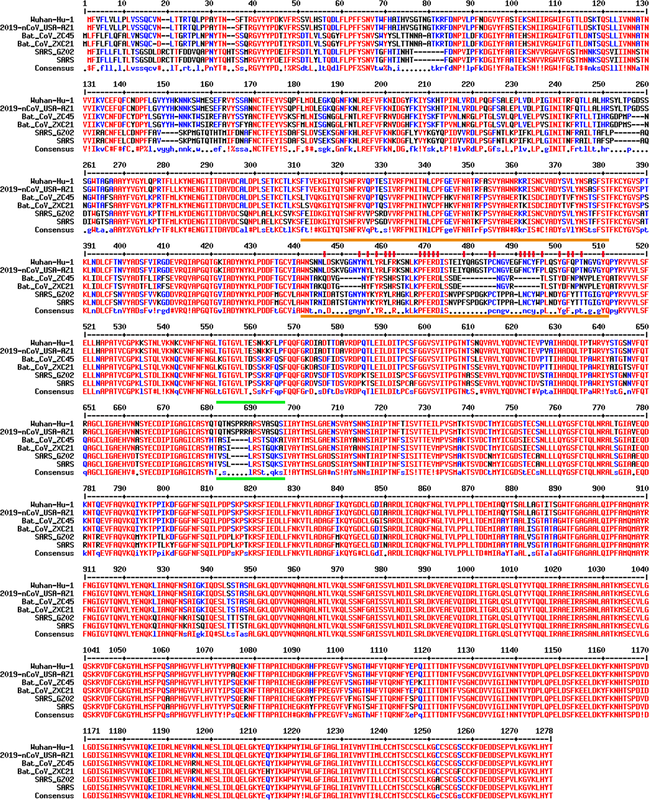
?圖3. 相關冠狀病毒的spike蛋白的序列比對。參與比對的包括兩個是武漢冠狀病毒(Wuhan-Hu-1, 2019-nCoV_USA-AZ1),舟山蝙蝠病毒(Bat_CoV_ZC45,Bat_CoV_ZXC21), 和SARS冠狀病毒(SARS_GZ02,SARS)。兩條橙色線之間的是和ACE2結合的關鍵區間。其中對結合起至關重要作用的氨基酸以紅色豎杠標記。兩條綠色線之間的是furin酶切位點。這樣的位點隻存在於武漢冠狀病毒,而在其他任何的beta類冠狀病毒中從來沒有觀察到過。序列對比用的是MultAlin webserver (http://multalin.toulouse.inra.fr/multalin/)。
圖3是六個不同冠狀病毒的spike蛋白的序列比對。其中,兩個是武漢冠狀病毒(Wuhan-Hu-1, 2019-nCoV_USA-AZ1);兩個是舟山蝙蝠病毒(Bat_CoV_ZC45,Bat_CoV_ZXC21);兩個是SARS冠狀病毒(SARS_GZ02,SARS)。快速掃一眼,你就能發現對於spike蛋白的後半部分(690之後),也就是S2部分,六個病毒的序列都很接近。區別主要在前半部分(1-~690),也就是S1。假如你觀察位於最上麵的四個病毒的話,即兩個武漢冠狀病毒和兩個舟山蝙蝠病毒,你會發現它們的S1也大體一致,唯有兩三處明顯不同。然而,就是這幾處不同,尤其是不同裏麵的細節,基本上讓我和很多其他人確信,這個武漢冠狀病毒是人造的,而絕非來源於大自然。
第一個重要的不同之處就是圖中用橙色線標注的地方。很明顯,武漢冠狀病毒在這一個區間內與舟山蝙蝠病毒非常不一樣。與之形成反襯的是,在整個基因組的其他任何區域兩種病毒都是高度一致的(超過 95% 的一致性)。與此同時,武漢冠狀病毒的這一段卻與SARS的spike蛋白的相關片段高度相似。注意,這個用橙色線標注出來的部分就是圖2C中顯示為橙色的肽段。像我們之前提到的那樣,這部分肽段包括了所有和ACE2結合所需要的微觀結構。一個很直觀的感覺就是,SARS中這個最關鍵的片段被“複製”了,然後“粘貼”進了舟山蝙蝠病毒中,從而製造出了武漢冠狀病毒。
再稍微仔細一點兒觀察後,你可能會發現武漢冠狀病毒和SARS冠狀病毒在這一段似乎還有相異之處,因此又不是很明顯的“複製”和“粘貼”。可是,當你非常仔細地在微觀結構層麵分析SARS的spike蛋白如何與人體ACE2蛋白結合之後(圖2C),你會發現S1中所有對結合ACE2至關重要的氨基酸(圖2C中以橙色棍狀標注的,同時也是圖3中用紅色豎杠標注的氨基酸)都基本被精確保留了或者是替換成了性質類似的氨基酸。另一方麵,兩種病毒的spike蛋白在這一區間的不同之處基本全在不影響與ACE2結合的位置上。僅從序列上的這種信息就可以判斷,武漢冠狀病毒不僅能與ACE2結合,而且其結合的方式與SARS的spike蛋白結合ACE2的方式決無二致(圖2BC)。
而舟山蝙蝠病毒,因為它們的spike蛋白缺少很多關鍵的氨基酸,它們與ACE2結合進而侵染人體的可能性是非常非常小的。
我們再梳理一遍武漢冠狀病毒和舟山蝙蝠病毒的序列比較:兩者在絕大部分蛋白上都幾乎一模一樣(95%一致性),而唯獨在spike蛋白的S1這裏,一段SARS的spike蛋白中專門決定與人體ACE2結合的序列被生生的“繼承”了過來,替換掉了的是原舟山蝙蝠病毒中一段無法與人體ACE2結合的肽段。而且這個還並不是生搬硬套的繼承:所有的關鍵氨基酸被保留而不參與結合ACE2的很多氨基酸都變了。這種方式的“替換”不可謂不神奇,而這個神奇“替換”的結果就是武漢冠狀病毒“獲得”了感染人的能力。此能力是舟山蝙蝠病毒所不具備的。
自然進化能如此精準,同時具有如此的隱蔽性嗎???
假如以上還不算觸目驚心的話,我們再看看這裏麵的第二個“魔術”。這次關注的焦點是圖3中兩條綠線之間的部分。在這裏,唯有武漢冠狀病毒有一小段額外的序列,就是SPRRA。有了這個獨特的序列後,武漢冠狀病毒的spike蛋白就能夠在這個位置被人體的furin蛋白酶剪切。而此種剪切是被證明可以增強流感病毒(含有類似的spike蛋白)的感染力的。需要注意的一點是,除了武漢冠狀病毒病毒之外,自然界中還沒有發現任何別的beta類冠狀病毒含有這樣的furin酶切位點。換句話說,還沒有任何證據表明,大自然以天然進化的方式產生過帶有furin酶切位點的beta類冠狀病毒。
進一步解釋為什麽這些序列上的變化不可能來自自然進化
我們之前簡短地解釋了為什麽隨機突變不可能導致一個類似舟山蝙蝠病毒的祖先進化成為武漢冠狀病毒。現在我們再進一步地說明這一點。確實,相對於其他部位來說,冠狀病毒的spike蛋白更容易出現變異。然而,當兩個病毒的S1差異很大時,必然是因為它們在很早很早之前就分開進化了,並且進化過程中頻繁地隨機變異以適應不同的宿主(因為S1是決定侵染什麽宿主的)。這種情況下,不光是兩種病毒的S1部分一致性低,它們整體序列一致性一定也是偏低的。而現在的情況是武漢冠狀病毒和舟山蝙蝠病毒全序列一致性高達95%,顯示這兩個病毒是非常近的近親。在這個前提下,出現S1蛋白一致性隻有69%的現象是完全不可思議的。所以說,武漢冠狀病毒的S1蛋白絕對不可能是由舟山蝙蝠病毒的S1蛋白通過隨機變異的方式進化而來的。
好,那我們再來說一下為什麽武漢冠狀病毒也不可能以基因重組的方式進化而來。
基因重組是指一個長基因中的某段可以被其他基因中與之類似的一段置換。基因重組也是自然進化的方式之一。它發生的頻率遠遠低於隨機突變。可它一旦發生,往往會給基因組裏的某些位置帶來非常大的,有時甚至很神奇的變化。
假如是自然進化中的基因重組導致了武漢冠狀病毒這個神奇的存在,它會怎麽發生呢?首先,這樣的基因重組要想發生必然是一個類似舟山蝙蝠病毒的祖先和另一個冠狀病毒同時侵染了一個動物體的同一個細胞。在這個前提下,非常偶然的會出現我們需要的基因重組,就是這個祖先病毒的某一段被另一個冠狀病毒中的相似片段所替換。非常重要的一點是,從一個舟山蝙蝠病毒那樣的祖先出發,最終進化出武漢冠狀病毒的話,一次基因重組是不夠的,必須有兩次基因重組才可能進化出如此的結果。其中一次,這個祖先病毒必須和一個類似SARS的冠狀病毒共存,然後通過基因重組精確地得到S1蛋白中專門和人體ACE2結合的那一小段(圖2和圖3中以橙色標記的部分)。除此之外,在另一個契機中,這個經過以上“改良”的祖先病毒還要再和一個在S1和S2之間包含furin酶切位點的冠狀病毒共存於一個細胞,並通過基因重組將這個furin酶切位點準確地置換了進來。
另外,像我們提過的那樣,基於舟山蝙蝠病毒和武漢冠狀病毒高度的一致性(95%),我們可以合理推測出兩種病毒並不是遠古之前分開進化的。也就是說,以上提到必須發生的兩個基因重組也都應當是相當近的年代發生的。
好,我們知道SARS的出現並感染人其實是進化中非常偶然的一件事。這也說明,要想自然中再出現一個和SARS類似,能侵染人,既結合人體ACE2,同時S1裏的關鍵序列又和SARS不太一樣的冠狀病毒應該是更偶然的一件事。再讓這樣的一個類SARS病毒與舟山蝙蝠病毒共存並發生精確的基因重組,那幾乎就是不可能的。除此以外,furin酶切位點在自然界中任何已知的beta類冠狀病毒的spike蛋白中都是不存在的。也就是說,讓舟山蝙蝠病毒和另外一個含有furin酶切位點的冠狀病毒共存於一個細胞,並通過基因重組將furin酶切位點置換進自己的基因組也是基本不可能的。
那麽好,讓這兩個幾乎不可能的事都發生的可能性是多少?我的答案是沒門兒,絕無可能。因此,武漢冠狀病毒絕對不可能來自於自然。
為什麽有些文獻必須被排除在外
有些關注了最近發表的科學論文的人可能會說,以上的分析是不對的,因為某個非常重要的發現並沒有被考慮進來。而恰巧,這個發現是與我的論點相悖,支持病毒來自自然進化的。那我為什麽敢對它完全無視呢?
簡單一點兒的答案就是,我認為這個發現, 或者說“證據”,很有可能是偽造的。
請允許我現在轉換我的角色,從一個科學家變成一個警探或者法官。假如我們把這個事件當成一個正在被調查或庭審的案件的話,那我們這裏有一個最大的嫌疑人,就是來自武漢病毒所和P4實驗室的冠狀病毒專家,石正麗博士。從武漢疫情爆發剛一開始,就不停的有人懷疑是石正麗人工製造了這個病毒,並有意或無意地把它釋放了出去。石一直是最大的,甚至唯一的嫌疑人。有意思的是,石在幾周前在自然雜誌(nature)發表了一篇論文(3)。裏麵她將當時剛剛公開不久的武漢冠狀病毒的序列與其它beta類冠狀病毒的序列做了比較,並由此描繪了武漢冠狀病毒的可能進化路徑。文章裏報道了一個全新的蝙蝠的冠狀病毒,RaTG13。此RaTG13與武漢冠狀病毒有非常之高的一致性。這裏最神奇的是,這個高一致性(98.5%)或者說非常低的變異率是均勻遍布於基因組的各個部分或者說各個蛋白的。我們之前懷疑武漢冠狀病毒來源於人造,其主要根據是武漢冠狀病毒與舟山蝙蝠病毒之間序列的一致性分布非常詭異,無法用自然進化解釋。這個問題在RaTG13身上則完全消失了。RaTG13的spike蛋白和武漢冠狀病毒的spike蛋白也高度一致(97.4%),與兩個病毒間在其他部位的“變異頻率”是一樣水平的。第一眼看過去,這個RaTG13很明顯就是與武漢冠狀病毒同源的,兩者不久之前有一個共同的祖先,從那分支並各自進化而來。這一“發現”有力地證明武漢冠狀病毒是來源於自然的。這篇報道了RaTG13的論文就是被我完全無視,在之前的分析中故意忽略的“證據”。
根據可靠的消息來源,石正麗跟幾個人分別承認過,她手中並沒有真正的RaTG13的毒株。據她說,她的實驗室是在七年前從雲南的蝙蝠的糞便中分析基因片段,從而尋找可能存在的冠狀病毒時發現的這個RaTG13的基因。用稍微通俗點兒的話說,事實上她是沒有實物證據能證明RaTG13存在的,她隻有這個病毒的基因序列,也就是一個由ATCG四個字母以各種方式組合出來的長鏈。
這個序列可不可能被偽造呢?這個其實再容易不過了。讓一個人坐那兒往一個word文檔裏打字就可以了。一共不到3萬個字母,不到一天就完活兒了。要是你還有一個模版序列,它和你想要得到的序列有98%是一摸一樣的,那這個工作就要更容易一千倍了。新序列打完之後,隻要上傳到網上公開的基因數據庫就可以了,期間沒有什麽嚴格的審核。上傳了的序列就可以被公開引用,並拿來分析數據和發表文章了。
那麽,現在的問題就是,這樣一個RaTG13的序列能不能拿來當作證據呢?別忘了,這個事很核心的一點就是到底是不是石正麗本人製造了這個病毒。假如真是人為製造的話,那麽這可以說是人類有史以來最大的犯罪,而石是這裏唯一最大的嫌疑人。這種情況下,石是不是有為自己掩蓋罪行的動機呢?假如她在自證清白時所用的證據不過是她剛剛在word文檔中打出來的一連串字母,法官,陪審團,受害者,或者任何人應不應該認可這樣的證據呢?
非常不幸的是,最近發表的很多科研論文都把分析和推理建立在這個RaTG13的序列之上。它們的結論也自然毫無意外:武漢冠狀病毒一定來源於自然進化。我希望讀者們能認識到或認同我的看法,就是這個RaTG13的序列很可能是石正麗偽造的,所以以此為依據得到的任何所謂科學判斷都需要打個大大的問號。
好,我們現在再換一個角度看。這個RaTG13病毒的序列是很震撼的。任何這個領域的專家隻要瞥一眼它的spike蛋白的序列就會立馬意識到這個病毒和SARS很像,很可能可以結合人體ACE2,所以它非常可能具有感染人的能力。石本人就是這樣的專家。石正麗實驗室對外宣稱,她們研究的目的就是希望通過對蝙蝠的冠狀病毒的充分了解達到能夠預警人類的作用,防止類似SARS那樣的健康災難的發生。那麽本著這樣的心態,石正麗怎麽會在七年之間似乎完全忽視了一個像RaTG13這樣的病毒呢?她怎麽能忍七年而不發表這個驚人的發現呢?為什麽隻是在武漢疫情爆發之後,當人們開始懷疑武漢病毒的來源的時候,石突然決定發表這個RaTG13的序列呢?
所有這些都不符合常理。這些事實放在一塊兒隻能讓人更加地懷疑石正麗。她或者直接參與了製造這個病毒,或者在幫忙掩蓋真相,或是兩者兼而有之。另外,這些事實當然也更加說明,這個RaTG13的序列必須被排除於任何的科學分析之中。
同樣需要排除在外的是所謂穿山甲是病毒從蝙蝠傳播到人的中間宿主這一說法。二月初的時候,華南農業大學的三位研究人員召開了一個新聞發布會,聲稱他們最新的科研結果顯示穿山甲很可能是病毒的中間宿主。首先,這個新聞發布會召開的時間點就很可疑,當時坊間開始討論說蝙蝠病毒不可能直接傳染人,需要有一個中間宿主(因為病毒的spike蛋白需要在一個有類似於人的ACE2的動物中“學習”和適應如何與這樣的ACE2結合)。以往的經驗告訴我們,當什麽事情必須存在才能讓中共自圓其說的時候,這個事情一定會神奇的出現或者被某個中共控製的人“親口證實”,就像石正麗的RaTG13。這次從天而降的是穿山甲的冠狀病毒(從而佐證穿山甲是中間宿主)。這些科研人員都等不到發文章了,直接表示他們有證據:一個序列(又是序列)。具體來說,是他們發現的穿山甲冠狀病毒的S1蛋白的受體結合區域(也就是和ACE2結合的區域,英文簡稱RBD)的序列信息。和之前一樣,沒有活體病毒,隻是序列信息,而且當時還沒公布。像我們在討論RaTG13時提到的,這樣的序列信息在word文檔裏打字就可以了。這次這個RBD更短,幾分鍾就能打完了。基於這種情況,這個穿山甲冠狀病毒的序列信息同樣有很大的可能性是中共為混淆視聽而偽造的。
幸運的是,領域裏人似乎也很快就放過了穿山甲。最終這些穿山甲冠狀病毒的序列信息被兩個科研組(華南農大和香港的管軼的科研組)分別發表並公布出來(4,5),可是這些序列卻沒能讓業界相信穿山甲是中間宿主。這其中有一部分原因是穿山甲冠狀病毒的spike蛋白序列中也沒觀察到furin酶切位點。
所以,就像RaTG13一樣,最近這些證實穿山甲為中間宿主的科研論文也應該在分析時被排除在外。事實上,就在剛剛,這幾個華南農大的研究人員向媒體承認,在對穿山甲冠狀病毒的全序列分析之後,他們認為自己之前的說法有誤,穿山甲不是武漢冠狀病毒可能的中間宿主。
一些應該被高度關注的科研文獻
我們剛剛花大篇幅解釋了為什麽某些“科研證據”必須被排除在外。現在我們再來看看某些其他的文獻和證據必須被高度關注和認真參考。
第一,這兩個與武漢冠狀病毒具有詭異相似性的舟山蝙蝠病毒(ZC45和ZXC21)是由中共的軍事科研單位收集並發現的。他們把科研結果,其中包括舟山蝙蝠病毒的序列信息,在2018年發表了(6)。在這裏我想強調兩點:一,如果武漢冠狀病毒是人造的,那它一定是用舟山蝙蝠病毒為模版(template)做的;二,這篇文章揭示全世界隻有中共手中有舟山蝙蝠病毒。
第二,石正麗在2015年與人合作發表了一篇Nature Medicine的文章(7)。他的合作者是北卡大學的Ralph Baric教授。這篇文章揭示了如何把一個普通的蝙蝠冠狀病毒變成一個能侵染人的病毒。具體來說,他們把一個原本對人無感染能力的蝙蝠冠狀病毒的spike蛋白換成了能夠與人體ACE2結合的一個spike蛋白。這一個改變就把原來的蝙蝠病毒變成了針對人的冠狀病毒。好,現在我們回頭看看,在武漢冠狀病毒發生的事基本就是按著這篇文章的腳本兒來的。現實中,並不是整個的spike蛋白被替換了,而隻是替換了spike蛋白的S1中最關鍵的一部分。這個最小程度的改變卻充分地給了被改造後的病毒感染人的能力。這裏真正的不同之處是,這種微妙地改變spike蛋白的方式讓試圖找尋病毒來源的人們更容易迷失方向。
第三,一篇2006的文獻揭示,以人為的方式在SARS病毒spike的S1和S2之間插入一個furin酶切位點之後,病毒的傳染性如預測的一樣出現了大幅度的提高(8)。神奇的是,這個操作也如出一轍地出現在了武漢冠狀病毒的spike蛋白之中(圖3中綠線標注的肽段)。而且,這篇文章還推斷,由於具有更高的感染性,如此改造之後的SARS冠狀病毒很可能會進一步感染肺以外的其它器官。假如你關注了一些最近有關武漢冠狀病毒的病症特征的話,你可能已經注意到了,幾個獨立的報道已經描述了這個病毒不但攻擊肺,同時也能攻擊肝,心,血管,甚至中樞神經係統等等。
清晰的證據鏈和邏輯鏈顯示中共為製造武漢冠狀病毒的元凶
假如你把以上這些信息稍作整理,你會發現假如是中共製造了武漢冠狀病毒的話,這件事是多麽的符合邏輯。很明顯,製造這個病毒所用的的模版,也就是舟山蝙蝠病毒,隻有中共手裏才有(6)。在這個基礎上,他們要做的就是改變這個蝙蝠病毒的某些地方,讓它變得能感染人,而且最好是傳染性極強。而實際上發生的就是他們完全按著兩個發表過的概念做出了這個新病毒(7,8):1)他們依照SARS蛋白結合ACE2的方式對舟山蝙蝠病毒的spike蛋白做了精確的改造,使之能按照SARS的spike那樣的方式結合人體ACE2。如此一來,新病毒變得能感染人了;2)他們在spike蛋白的S1和S2之間插入了一個可以讓furin酶來切割的位點,從而讓病毒增加了幾倍甚至幾十倍的感染力。這兩個概念可以說是中外文獻中能找到的,僅有的兩個如何改進冠狀病毒的方式。結果,兩者在武漢冠狀病毒的身上都被完美的體現了。假如是大自然製造了這個病毒,那大自然一定非常仔細地研究了近些年人類在這方麵的科研成果,並且把人類的幾個最重要的發現極為忠實地應用在了她的工作之中(2,6,7,8)。
我們也可以再往回想一想。為什麽中共治下有那麽多的科研單位這麽努力地在各處尋找冠狀病毒呢?事實真是像他們說的那樣,是為了更好地了解冠狀病毒各種可能性,進而更好地預測未來可能由新的冠狀病毒導致的疫情嗎?那他們為什麽不把精力花在製造廣譜的冠狀病毒疫苗或者是針對冠狀病毒中的某個保守性高的藥物靶點而製藥呢?後者不但對人類健康更有直接意義,而且比預測哪個冠狀病毒未來會爆發要容易不知多少倍。
另外一種可能性,就是他們收集這些野生動物的冠狀病毒是為了製造生物武器。到底哪一個是真相?你可以自己判斷。
對我來說,答案再明顯不過,這次的武漢冠狀病毒就是中共所製造的生物武器。
基於以上羅列的事實和對這些事實的分析,一個最合理的結論就是,除非中共能提供有效的證據證明自己是清白的,否則全世界已經有足夠的理由相信武漢冠狀病毒就是中共人工製造出來的。
引文出處:
1. Daniel Wrapp NW, Kizzmekia S. Corbett, Jory A. Goldsmith, Ching-Lin Hsieh, Olubukola Abiona, Barney S. Graham, Jason S. McLellan. Cryo-EM Structure of the 2019-nCoV Spike in the Prefusion Conformation. Science. 2020.
2. Song W, Gui M, Wang X, Xiang Y. Cryo-EM structure of the SARS coronavirus spike glycoprotein in complex with its host cell receptor ACE2. PLoS Pathog. 2018;14(8):e1007236.
3. Zhou P, Yang XL, Wang XG, Hu B, Zhang L, Zhang W, et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature. 2020.
4. Kangpeng Xiao JZ, Yaoyu Feng, Niu Zhou, Xu Zhang, Jie-Jian Zou, Na Li, Yaqiong Guo, Xiaobing Li, Xuejuan Shen, Zhipeng Zhang, Fanfan Shu, Wanyi Huang, Yu Li, Ziding Zhang, Rui-Ai Chen, Ya-Jiang Wu, Shi-Ming Peng, Mian Huang, Wei-Jun Xie, Qin-Hui Cai, Fang-Hui Hou, Yahong Liu, Wu Chen, Lihua Xiao, Yongyi Shen. Isolation and Characterization of 2019-nCoV-like Coronavirus from Malayan Pangolins. bioRxiv. 2020.
5. Tommy Tsan-Yuk Lam MH-HS, Hua-Chen Zhu, Yi-Gang Tong, Xue-Bing Ni, Yun-Shi Liao, Wei Wei, William Yiu-Man Cheung, Wen-Juan Li, Lian-Feng Li, Gabriel M Leung, Edward C. Holmes, Yan-Ling Hu, Yi Guan. Identification of 2019-nCoV related coronaviruses in Malayan pangolins in southern China. bioRxiv. 2020.
6. Hu D, Zhu C, Ai L, He T, Wang Y, Ye F, et al. Genomic characterization and infectivity of a novel SARS-like coronavirus in Chinese bats. Emerg Microbes Infect. 2018;7(1):154.
7. Menachery VD, Yount BL, Jr., Debbink K, Agnihothram S, Gralinski LE, Plante JA, et al. A SARS-like cluster of circulating bat coronaviruses shows potential for human emergence. Nat Med. 2015;21(12):1508-13.
8. Follis KE, York J, Nunberg JH. Furin cleavage of the SARS coronavirus spike glycoprotein enhances cell-cell fusion but does not affect virion entry. Virology. 2006;350(2):358-69.
圖3是六個不同冠狀病毒的spike蛋白的序列比對。其中,兩個是武漢冠狀病毒(Wuhan-Hu-1, 2019-nCoV_USA-AZ1);兩個是舟山蝙蝠病毒(Bat_CoV_ZC45,Bat_CoV_ZXC21);兩個是SARS冠狀病毒(SARS_GZ02,SARS)。快速掃一眼,你就能發現對於spike蛋白的後半部分(690之後),也就是S2部分,六個病毒的序列都很接近。區別主要在前半部分(1-~690),也就是S1。假如你觀察位於最上麵的四個病毒的話,即兩個武漢冠狀病毒和兩個舟山蝙蝠病毒,你會發現它們的S1也大體一致,唯有兩三處明顯不同。然而,就是這幾處不同,尤其是不同裏麵的細節,基本上讓我和很多其他人確信,這個武漢冠狀病毒是人造的,而絕非來源於大自然。
第一個重要的不同之處就是圖中用橙色線標注的地方。很明顯,武漢冠狀病毒在這一個區間內與舟山蝙蝠病毒非常不一樣。與之形成反襯的是,在整個基因組的其他任何區域兩種病毒都是高度一致的(超過 95% 的一致性)。與此同時,武漢冠狀病毒的這一段卻與SARS的spike蛋白的相關片段高度相似。注意,這個用橙色線標注出來的部分就是圖2C中顯示為橙色的肽段。像我們之前提到的那樣,這部分肽段包括了所有和ACE2結合所需要的微觀結構。一個很直觀的感覺就是,SARS中這個最關鍵的片段被“複製”了,然後“粘貼”進了舟山蝙蝠病毒中,從而製造出了武漢冠狀病毒。
再稍微仔細一點兒觀察後,你可能會發現武漢冠狀病毒和SARS冠狀病毒在這一段似乎還有相異之處,因此又不是很明顯的“複製”和“粘貼”。可是,當你非常仔細地在微觀結構層麵分析SARS的spike蛋白如何與人體ACE2蛋白結合之後(圖2C),你會發現S1中所有對結合ACE2至關重要的氨基酸(圖2C中以橙色棍狀標注的,同時也是圖3中用紅色豎杠標注的氨基酸)都基本被精確保留了或者是替換成了性質類似的氨基酸。另一方麵,兩種病毒的spike蛋白在這一區間的不同之處基本全在不影響與ACE2結合的位置上。僅從序列上的這種信息就可以判斷,武漢冠狀病毒不僅能與ACE2結合,而且其結合的方式與SARS的spike蛋白結合ACE2的方式決無二致(圖2BC)。
而舟山蝙蝠病毒,因為它們的spike蛋白缺少很多關鍵的氨基酸,它們與ACE2結合進而侵染人體的可能性是非常非常小的。
我們再梳理一遍武漢冠狀病毒和舟山蝙蝠病毒的序列比較:兩者在絕大部分蛋白上都幾乎一模一樣(95%一致性),而唯獨在spike蛋白的S1這裏,一段SARS的spike蛋白中專門決定與人體ACE2結合的序列被生生的“繼承”了過來,替換掉了的是原舟山蝙蝠病毒中一段無法與人體ACE2結合的肽段。而且這個還並不是生搬硬套的繼承:所有的關鍵氨基酸被保留而不參與結合ACE2的很多氨基酸都變了。這種方式的“替換”不可謂不神奇,而這個神奇“替換”的結果就是武漢冠狀病毒“獲得”了感染人的能力。此能力是舟山蝙蝠病毒所不具備的。
自然進化能如此精準,同時具有如此的隱蔽性嗎???
假如以上還不算觸目驚心的話,我們再看看這裏麵的第二個“魔術”。這次關注的焦點是圖3中兩條綠線之間的部分。在這裏,唯有武漢冠狀病毒有一小段額外的序列,就是SPRRA。有了這個獨特的序列後,武漢冠狀病毒的spike蛋白就能夠在這個位置被人體的furin蛋白酶剪切。而此種剪切是被證明可以增強流感病毒(含有類似的spike蛋白)的感染力的。需要注意的一點是,除了武漢冠狀病毒病毒之外,自然界中還沒有發現任何別的beta類冠狀病毒含有這樣的furin酶切位點。換句話說,還沒有任何證據表明,大自然以天然進化的方式產生過帶有furin酶切位點的beta類冠狀病毒。
進一步解釋為什麽這些序列上的變化不可能來自自然進化
我們之前簡短地解釋了為什麽隨機突變不可能導致一個類似舟山蝙蝠病毒的祖先進化成為武漢冠狀病毒。現在我們再進一步地說明這一點。確實,相對於其他部位來說,冠狀病毒的spike蛋白更容易出現變異。然而,當兩個病毒的S1差異很大時,必然是因為它們在很早很早之前就分開進化了,並且進化過程中頻繁地隨機變異以適應不同的宿主(因為S1是決定侵染什麽宿主的)。這種情況下,不光是兩種病毒的S1部分一致性低,它們整體序列一致性一定也是偏低的。而現在的情況是武漢冠狀病毒和舟山蝙蝠病毒全序列一致性高達95%,顯示這兩個病毒是非常近的近親。在這個前提下,出現S1蛋白一致性隻有69%的現象是完全不可思議的。所以說,武漢冠狀病毒的S1蛋白絕對不可能是由舟山蝙蝠病毒的S1蛋白通過隨機變異的方式進化而來的。
好,那我們再來說一下為什麽武漢冠狀病毒也不可能以基因重組的方式進化而來。
基因重組是指一個長基因中的某段可以被其他基因中與之類似的一段置換。基因重組也是自然進化的方式之一。它發生的頻率遠遠低於隨機突變。可它一旦發生,往往會給基因組裏的某些位置帶來非常大的,有時甚至很神奇的變化。
假如是自然進化中的基因重組導致了武漢冠狀病毒這個神奇的存在,它會怎麽發生呢?首先,這樣的基因重組要想發生必然是一個類似舟山蝙蝠病毒的祖先和另一個冠狀病毒同時侵染了一個動物體的同一個細胞。在這個前提下,非常偶然的會出現我們需要的基因重組,就是這個祖先病毒的某一段被另一個冠狀病毒中的相似片段所替換。非常重要的一點是,從一個舟山蝙蝠病毒那樣的祖先出發,最終進化出武漢冠狀病毒的話,一次基因重組是不夠的,必須有兩次基因重組才可能進化出如此的結果。其中一次,這個祖先病毒必須和一個類似SARS的冠狀病毒共存,然後通過基因重組精確地得到S1蛋白中專門和人體ACE2結合的那一小段(圖2和圖3中以橙色標記的部分)。除此之外,在另一個契機中,這個經過以上“改良”的祖先病毒還要再和一個在S1和S2之間包含furin酶切位點的冠狀病毒共存於一個細胞,並通過基因重組將這個furin酶切位點準確地置換了進來。
另外,像我們提過的那樣,基於舟山蝙蝠病毒和武漢冠狀病毒高度的一致性(95%),我們可以合理推測出兩種病毒並不是遠古之前分開進化的。也就是說,以上提到必須發生的兩個基因重組也都應當是相當近的年代發生的。
好,我們知道SARS的出現並感染人其實是進化中非常偶然的一件事。這也說明,要想自然中再出現一個和SARS類似,能侵染人,既結合人體ACE2,同時S1裏的關鍵序列又和SARS不太一樣的冠狀病毒應該是更偶然的一件事。再讓這樣的一個類SARS病毒與舟山蝙蝠病毒共存並發生精確的基因重組,那幾乎就是不可能的。除此以外,furin酶切位點在自然界中任何已知的beta類冠狀病毒的spike蛋白中都是不存在的。也就是說,讓舟山蝙蝠病毒和另外一個含有furin酶切位點的冠狀病毒共存於一個細胞,並通過基因重組將furin酶切位點置換進自己的基因組也是基本不可能的。
那麽好,讓這兩個幾乎不可能的事都發生的可能性是多少?我的答案是沒門兒,絕無可能。因此,武漢冠狀病毒絕對不可能來自於自然。
為什麽有些文獻必須被排除在外
有些關注了最近發表的科學論文的人可能會說,以上的分析是不對的,因為某個非常重要的發現並沒有被考慮進來。而恰巧,這個發現是與我的論點相悖,支持病毒來自自然進化的。那我為什麽敢對它完全無視呢?
簡單一點兒的答案就是,我認為這個發現, 或者說“證據”,很有可能是偽造的。
請允許我現在轉換我的角色,從一個科學家變成一個警探或者法官。假如我們把這個事件當成一個正在被調查或庭審的案件的話,那我們這裏有一個最大的嫌疑人,就是來自武漢病毒所和P4實驗室的冠狀病毒專家,石正麗博士。從武漢疫情爆發剛一開始,就不停的有人懷疑是石正麗人工製造了這個病毒,並有意或無意地把它釋放了出去。石一直是最大的,甚至唯一的嫌疑人。有意思的是,石在幾周前在自然雜誌(nature)發表了一篇論文(3)。裏麵她將當時剛剛公開不久的武漢冠狀病毒的序列與其它beta類冠狀病毒的序列做了比較,並由此描繪了武漢冠狀病毒的可能進化路徑。文章裏報道了一個全新的蝙蝠的冠狀病毒,RaTG13。此RaTG13與武漢冠狀病毒有非常之高的一致性。這裏最神奇的是,這個高一致性(98.5%)或者說非常低的變異率是均勻遍布於基因組的各個部分或者說各個蛋白的。我們之前懷疑武漢冠狀病毒來源於人造,其主要根據是武漢冠狀病毒與舟山蝙蝠病毒之間序列的一致性分布非常詭異,無法用自然進化解釋。這個問題在RaTG13身上則完全消失了。RaTG13的spike蛋白和武漢冠狀病毒的spike蛋白也高度一致(97.4%),與兩個病毒間在其他部位的“變異頻率”是一樣水平的。第一眼看過去,這個RaTG13很明顯就是與武漢冠狀病毒同源的,兩者不久之前有一個共同的祖先,從那分支並各自進化而來。這一“發現”有力地證明武漢冠狀病毒是來源於自然的。這篇報道了RaTG13的論文就是被我完全無視,在之前的分析中故意忽略的“證據”。
根據可靠的消息來源,石正麗跟幾個人分別承認過,她手中並沒有真正的RaTG13的毒株。據她說,她的實驗室是在七年前從雲南的蝙蝠的糞便中分析基因片段,從而尋找可能存在的冠狀病毒時發現的這個RaTG13的基因。用稍微通俗點兒的話說,事實上她是沒有實物證據能證明RaTG13存在的,她隻有這個病毒的基因序列,也就是一個由ATCG四個字母以各種方式組合出來的長鏈。
這個序列可不可能被偽造呢?這個其實再容易不過了。讓一個人坐那兒往一個word文檔裏打字就可以了。一共不到3萬個字母,不到一天就完活兒了。要是你還有一個模版序列,它和你想要得到的序列有98%是一摸一樣的,那這個工作就要更容易一千倍了。新序列打完之後,隻要上傳到網上公開的基因數據庫就可以了,期間沒有什麽嚴格的審核。上傳了的序列就可以被公開引用,並拿來分析數據和發表文章了。
那麽,現在的問題就是,這樣一個RaTG13的序列能不能拿來當作證據呢?別忘了,這個事很核心的一點就是到底是不是石正麗本人製造了這個病毒。假如真是人為製造的話,那麽這可以說是人類有史以來最大的犯罪,而石是這裏唯一最大的嫌疑人。這種情況下,石是不是有為自己掩蓋罪行的動機呢?假如她在自證清白時所用的證據不過是她剛剛在word文檔中打出來的一連串字母,法官,陪審團,受害者,或者任何人應不應該認可這樣的證據呢?
非常不幸的是,最近發表的很多科研論文都把分析和推理建立在這個RaTG13的序列之上。它們的結論也自然毫無意外:武漢冠狀病毒一定來源於自然進化。我希望讀者們能認識到或認同我的看法,就是這個RaTG13的序列很可能是石正麗偽造的,所以以此為依據得到的任何所謂科學判斷都需要打個大大的問號。
好,我們現在再換一個角度看。這個RaTG13病毒的序列是很震撼的。任何這個領域的專家隻要瞥一眼它的spike蛋白的序列就會立馬意識到這個病毒和SARS很像,很可能可以結合人體ACE2,所以它非常可能具有感染人的能力。石本人就是這樣的專家。石正麗實驗室對外宣稱,她們研究的目的就是希望通過對蝙蝠的冠狀病毒的充分了解達到能夠預警人類的作用,防止類似SARS那樣的健康災難的發生。那麽本著這樣的心態,石正麗怎麽會在七年之間似乎完全忽視了一個像RaTG13這樣的病毒呢?她怎麽能忍七年而不發表這個驚人的發現呢?為什麽隻是在武漢疫情爆發之後,當人們開始懷疑武漢病毒的來源的時候,石突然決定發表這個RaTG13的序列呢?
所有這些都不符合常理。這些事實放在一塊兒隻能讓人更加地懷疑石正麗。她或者直接參與了製造這個病毒,或者在幫忙掩蓋真相,或是兩者兼而有之。另外,這些事實當然也更加說明,這個RaTG13的序列必須被排除於任何的科學分析之中。
同樣需要排除在外的是所謂穿山甲是病毒從蝙蝠傳播到人的中間宿主這一說法。二月初的時候,華南農業大學的三位研究人員召開了一個新聞發布會,聲稱他們最新的科研結果顯示穿山甲很可能是病毒的中間宿主。首先,這個新聞發布會召開的時間點就很可疑,當時坊間開始討論說蝙蝠病毒不可能直接傳染人,需要有一個中間宿主(因為病毒的spike蛋白需要在一個有類似於人的ACE2的動物中“學習”和適應如何與這樣的ACE2結合)。以往的經驗告訴我們,當什麽事情必須存在才能讓中共自圓其說的時候,這個事情一定會神奇的出現或者被某個中共控製的人“親口證實”,就像石正麗的RaTG13。這次從天而降的是穿山甲的冠狀病毒(從而佐證穿山甲是中間宿主)。這些科研人員都等不到發文章了,直接表示他們有證據:一個序列(又是序列)。具體來說,是他們發現的穿山甲冠狀病毒的S1蛋白的受體結合區域(也就是和ACE2結合的區域,英文簡稱RBD)的序列信息。和之前一樣,沒有活體病毒,隻是序列信息,而且當時還沒公布。像我們在討論RaTG13時提到的,這樣的序列信息在word文檔裏打字就可以了。這次這個RBD更短,幾分鍾就能打完了。基於這種情況,這個穿山甲冠狀病毒的序列信息同樣有很大的可能性是中共為混淆視聽而偽造的。
幸運的是,領域裏人似乎也很快就放過了穿山甲。最終這些穿山甲冠狀病毒的序列信息被兩個科研組(華南農大和香港的管軼的科研組)分別發表並公布出來(4,5),可是這些序列卻沒能讓業界相信穿山甲是中間宿主。這其中有一部分原因是穿山甲冠狀病毒的spike蛋白序列中也沒觀察到furin酶切位點。
所以,就像RaTG13一樣,最近這些證實穿山甲為中間宿主的科研論文也應該在分析時被排除在外。事實上,就在剛剛,這幾個華南農大的研究人員向媒體承認,在對穿山甲冠狀病毒的全序列分析之後,他們認為自己之前的說法有誤,穿山甲不是武漢冠狀病毒可能的中間宿主。
一些應該被高度關注的科研文獻
我們剛剛花大篇幅解釋了為什麽某些“科研證據”必須被排除在外。現在我們再來看看某些其他的文獻和證據必須被高度關注和認真參考。
第一,這兩個與武漢冠狀病毒具有詭異相似性的舟山蝙蝠病毒(ZC45和ZXC21)是由中共的軍事科研單位收集並發現的。他們把科研結果,其中包括舟山蝙蝠病毒的序列信息,在2018年發表了(6)。在這裏我想強調兩點:一,如果武漢冠狀病毒是人造的,那它一定是用舟山蝙蝠病毒為模版(template)做的;二,這篇文章揭示全世界隻有中共手中有舟山蝙蝠病毒。
第二,石正麗在2015年與人合作發表了一篇Nature Medicine的文章(7)。他的合作者是北卡大學的Ralph Baric教授。這篇文章揭示了如何把一個普通的蝙蝠冠狀病毒變成一個能侵染人的病毒。具體來說,他們把一個原本對人無感染能力的蝙蝠冠狀病毒的spike蛋白換成了能夠與人體ACE2結合的一個spike蛋白。這一個改變就把原來的蝙蝠病毒變成了針對人的冠狀病毒。好,現在我們回頭看看,在武漢冠狀病毒發生的事基本就是按著這篇文章的腳本兒來的。現實中,並不是整個的spike蛋白被替換了,而隻是替換了spike蛋白的S1中最關鍵的一部分。這個最小程度的改變卻充分地給了被改造後的病毒感染人的能力。這裏真正的不同之處是,這種微妙地改變spike蛋白的方式讓試圖找尋病毒來源的人們更容易迷失方向。
第三,一篇2006的文獻揭示,以人為的方式在SARS病毒spike的S1和S2之間插入一個furin酶切位點之後,病毒的傳染性如預測的一樣出現了大幅度的提高(8)。神奇的是,這個操作也如出一轍地出現在了武漢冠狀病毒的spike蛋白之中(圖3中綠線標注的肽段)。而且,這篇文章還推斷,由於具有更高的感染性,如此改造之後的SARS冠狀病毒很可能會進一步感染肺以外的其它器官。假如你關注了一些最近有關武漢冠狀病毒的病症特征的話,你可能已經注意到了,幾個獨立的報道已經描述了這個病毒不但攻擊肺,同時也能攻擊肝,心,血管,甚至中樞神經係統等等。
清晰的證據鏈和邏輯鏈顯示中共為製造武漢冠狀病毒的元凶
假如你把以上這些信息稍作整理,你會發現假如是中共製造了武漢冠狀病毒的話,這件事是多麽的符合邏輯。很明顯,製造這個病毒所用的的模版,也就是舟山蝙蝠病毒,隻有中共手裏才有(6)。在這個基礎上,他們要做的就是改變這個蝙蝠病毒的某些地方,讓它變得能感染人,而且最好是傳染性極強。而實際上發生的就是他們完全按著兩個發表過的概念做出了這個新病毒(7,8):1)他們依照SARS蛋白結合ACE2的方式對舟山蝙蝠病毒的spike蛋白做了精確的改造,使之能按照SARS的spike那樣的方式結合人體ACE2。如此一來,新病毒變得能感染人了;2)他們在spike蛋白的S1和S2之間插入了一個可以讓furin酶來切割的位點,從而讓病毒增加了幾倍甚至幾十倍的感染力。這兩個概念可以說是中外文獻中能找到的,僅有的兩個如何改進冠狀病毒的方式。結果,兩者在武漢冠狀病毒的身上都被完美的體現了。假如是大自然製造了這個病毒,那大自然一定非常仔細地研究了近些年人類在這方麵的科研成果,並且把人類的幾個最重要的發現極為忠實地應用在了她的工作之中(2,6,7,8)。
我們也可以再往回想一想。為什麽中共治下有那麽多的科研單位這麽努力地在各處尋找冠狀病毒呢?事實真是像他們說的那樣,是為了更好地了解冠狀病毒各種可能性,進而更好地預測未來可能由新的冠狀病毒導致的疫情嗎?那他們為什麽不把精力花在製造廣譜的冠狀病毒疫苗或者是針對冠狀病毒中的某個保守性高的藥物靶點而製藥呢?後者不但對人類健康更有直接意義,而且比預測哪個冠狀病毒未來會爆發要容易不知多少倍。
另外一種可能性,就是他們收集這些野生動物的冠狀病毒是為了製造生物武器。到底哪一個是真相?你可以自己判斷。
對我來說,答案再明顯不過,這次的武漢冠狀病毒就是中共所製造的生物武器。
基於以上羅列的事實和對這些事實的分析,一個最合理的結論就是,除非中共能提供有效的證據證明自己是清白的,否則全世界已經有足夠的理由相信武漢冠狀病毒就是中共人工製造出來的。
引文出處:
1. Daniel Wrapp NW, Kizzmekia S. Corbett, Jory A. Goldsmith, Ching-Lin Hsieh, Olubukola Abiona, Barney S. Graham, Jason S. McLellan. Cryo-EM Structure of the 2019-nCoV Spike in the Prefusion Conformation. Science. 2020.
2. Song W, Gui M, Wang X, Xiang Y. Cryo-EM structure of the SARS coronavirus spike glycoprotein in complex with its host cell receptor ACE2. PLoS Pathog. 2018;14(8):e1007236.
3. Zhou P, Yang XL, Wang XG, Hu B, Zhang L, Zhang W, et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature. 2020.
4. Kangpeng Xiao JZ, Yaoyu Feng, Niu Zhou, Xu Zhang, Jie-Jian Zou, Na Li, Yaqiong Guo, Xiaobing Li, Xuejuan Shen, Zhipeng Zhang, Fanfan Shu, Wanyi Huang, Yu Li, Ziding Zhang, Rui-Ai Chen, Ya-Jiang Wu, Shi-Ming Peng, Mian Huang, Wei-Jun Xie, Qin-Hui Cai, Fang-Hui Hou, Yahong Liu, Wu Chen, Lihua Xiao, Yongyi Shen. Isolation and Characterization of 2019-nCoV-like Coronavirus from Malayan Pangolins. bioRxiv. 2020.
5. Tommy Tsan-Yuk Lam MH-HS, Hua-Chen Zhu, Yi-Gang Tong, Xue-Bing Ni, Yun-Shi Liao, Wei Wei, William Yiu-Man Cheung, Wen-Juan Li, Lian-Feng Li, Gabriel M Leung, Edward C. Holmes, Yan-Ling Hu, Yi Guan. Identification of 2019-nCoV related coronaviruses in Malayan pangolins in southern China. bioRxiv. 2020.
6. Hu D, Zhu C, Ai L, He T, Wang Y, Ye F, et al. Genomic characterization and infectivity of a novel SARS-like coronavirus in Chinese bats. Emerg Microbes Infect. 2018;7(1):154.
7. Menachery VD, Yount BL, Jr., Debbink K, Agnihothram S, Gralinski LE, Plante JA, et al. A SARS-like cluster of circulating bat coronaviruses shows potential for human emergence. Nat Med. 2015;21(12):1508-13.
8. Follis KE, York J, Nunberg JH. Furin cleavage of the SARS coronavirus spike glycoprotein enhances cell-cell fusion but does not affect virion entry. Virology. 2006;350(2):358-69.

評論
目前還沒有任何評論
登錄後才可評論.